
Fosforilasi oksidatif adalah suatu lintasan metabolisme dengan penggunaan energi yang dilepaskan oleh oksidasi nutrien untuk menghasilkan ATP, dan mereduksi gas oksigen menjadi air.
Walaupun banyak bentuk kehidupan di bumi menggunakan berbagai jenis nutrien, hampir semua organisme menjalankan fosforilasi oksidatif untuk menghasilkan ATP, oleh karena efisiensi proses mendapatkan energi, dibandingkan dengan proses fermentasi alternatif lainnya seperti glikolisis anaerobik.
Menurut teori kemiosmotik yang dicetuskan oleh Peter Mitchell, energi yang dilepaskan dari reaksi oksidasi pada substrat pendonor elektron, baik pada respirasi aerobik maupun anaerobik, perlahan akan disimpan dalam bentuk potensial elektrokemis sepanjang garis tepi membran tempat terjadinya reaksi tersebut, yang kemudian dapat digunakan oleh ATP sintase untuk menginduksi reaksi fosforilasi terhadap molekul adenosina difosfat dengan molekul Pi.
Elektron yang melekat pada molekul sisi dalam kompleks IV rantai transpor elektron akan digunakan oleh kompleks V untuk menarik ion H+ dari sitoplasma menuju membran mitokondria sisi luar, disebut kopling kemiosmotik, yang menyebabkan kemiosmosis, yaitu difusi ion H+ melalui ATP sintase ke dalam mitokondria yang berlawanan dengan arah gradien pH, dari area dengan energi potensial elektrokimiawi lebih rendah menuju matriks dengan energi potensial lebih tinggi. Proses kopling kemiosmotik juga berpengaruh pada kombinasi gradien pH dan potensial listrik di sepanjang membran yang disebut gaya gerak proton.
Dari teori ini, keseluruhan reaksi kemudian disebut fosforilasi oksidatif.
Awal lintasan dimulai dari elektron yang dihasilkan oleh siklus asam sitrat yang ditransfer ke senyawa:
- NAD+ yang berada di dalam matriks mitokondria. Setelah menerima elektron, NAD+ akan bereaksi menjadi NADH dan ion H+, kemudian mendonorkan elektronnya ke rantai transpor elektron kompleks I.
- dan FAD yang berada di dalam rantai transpor elektron kompleks II. FAD akan menerima dua elektron, kemudian bereaksi menjadi FADH2 melalui reaksi redoks.
Walaupun fosforilasi oksidatif adalah bagian vital metabolisme, ia menghasilkan spesi oksigen reaktif seperti superoksida dan hidrogen peroksida pada kompleks I. Hal ini dapat mengakibatkan pembentukan radikal bebas, merusak sel tubuh, dan kemungkinan juga menyebabkan penuaan. Enzim-enzim yang terlibat dalam lintasan metabolisme ini juga merupakan target dari banyak obat dan racun yang dapat menghambat aktivitas enzim.
Tinjauan transfer energi melalui kemiosmosis
Fosforilasi oksidatif bekerja dengan cara menggunakan reaksi kimia yang menghasilkan energi untuk mendorong reaksi yang memerlukan energi. Kedua set reaksi ini dikatakan bergandengan. Hal ini berarti bahwa salah satu reaksi tidak dapat berjalan tanpa reaksi lainnya. Alur elektron melalui rantai transpor elektron adalah proses eksergonik, yakni melepaskan energi, manakala sintesis ATP adalah proses endergonik, yakni memerlukan energi. Baik rantai transpor elektron dan ATP sintase terdapat pada membran, dan energi ditransfer dari rantai transpor elektron ke ATP sintase melalui pergerakan proton melewati membran ini. Proses ini disebut sebagai kemiosmosis. Dalam praktiknya, ini mirip dengan sebuah sirkuit listrik, dengan arus proton didorong dari sisi negatif membran ke sisi positif oleh enzim pemompa proton yang ada pada rantai transpor elektron. Enzim ini seperti baterai. Pergerakan proton menciptakan gradien elektrokimiawi di sepanjang membran, yang sering disebut gaya gerak proton. Gradien ini mempunyai dua komponen: perbedaan pada konsentrasi proton (gradien pH) dan perbedaan pada potensi listrik. Energi tersimpan dalam bentuk perbedaan potensi listrik dalam mitokondria, dan juga sebagai gradien pH dalam kloroplas.
ATP sintase juga dapat memompa ion H+ keluar dari dalam matriks, apabila terjadi hidrolisis ATP pada kutub kompleksnya. Pada kasus hipertiroidisme pada hepatosit model tikus, juga ditemukan pemompaan ion H+ dari dalam matriks di luar mekanisme rantai transpor elektron, hal ini ditengarai terjadi oleh sebab peran hormon T3 yang dapat menyisip pada membran mitokondria sebelah dalam sebagai pompa ion.
Enzim ini seperti motor listrik, yang menggunakan gaya gerak proton untuk mendorong rotasi strukturnya dan menggunakan pergerakan ini untuk mensintesis ATP.
Energi yang dilepaskan oleh fosforilasi oksidatif ini cukup tinggi dibandingkan dengan energi yang dilepaskan oleh fermentasi anaerobik. Glikolisis hanya menghasilkan 2 molekul ATP, sedangkan pada fosforilasi oksidatif 10 molekul NADH dengan 2 molekul suksinat yang dibentuk dari konversi satu molekul glukosa menjadi karbon dioksida dan air, dihasilkan 30 sampai dengan 36 molekul ATP. Rendemen ATP ini sebenarnya merupakan nilai teoretis maksimum; pada praktiknya, ATP yang dihasilkan lebih rendah dari nilai tersebut.
Molekul pemindah elektron dan proton
Rantai transpor elektron membawa baik proton maupun elektron, mengangkut proton dari donor ke akseptor, dan mengangkut proton melawati membran. Proses ini menggunakan molekul yang larut dan terikat pada molekul transfer. Pada mitokondria, elektron ditransfer dalam ruang antarmembran menggunakan protein transfer elektron sitokrom c yang larut dalam air. Ia hanya mengangkut elektron, dan elektron ini ditransfer menggunakan reduksi dan oksidasi atom besi yang terikat pada protein pada gugus heme strukturnya. Sitokrom c juga ditemukan pada beberapa bakteri, di mana ia berlokasi di dalam ruang periplasma.
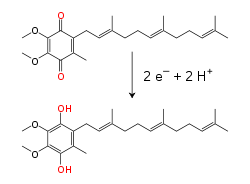
Dalam membran dalam mitokondria, koenzim Q10 pembawa elektron yang larut dalam lipid membawa baik elektron maupun proton menggunakan siklus redoks. Molekul benzokuinon yang kecil ini sangat hidrofobik, sehingga ia akan berdifusi dengan bebas ke dalam membran. Ketika Q menerima dua elektron dan dua proton, ia menjadi bentuk tereduksi ubikuinol (QH2); ketika QH2 melepaskan dua elektron dan dua proton, ia teroksidasi kembali menjadi bentuk ubikuinon (Q). Akibatnya, jika dua enzim disusun sedemikiannya Q direduksi pada satu sisi membran dan QH2 dioksidasi pada sisi lainnya, ubikuinon akan menggandengkan reaksi ini dan mengulang alik proton melewati membran. Beberapa rantai transpor elektron bakteri menggunakan kuinon yang berbeda, seperti menakuinon, selain ubikuinon.
Dalam protein, elektron ditransfer antar kofaktor flavin, gugus besi-sulfur, dan sitokrom. Terdapat beberapa jenis gugus besi-sulfur. Jenis paling sederhana yang ditemukan pada rantai transfer elektron terdiri dari dua atom besi yang dihubungkan oleh dua atom sulfur; ini disebut sebagai gugus [2Fe–2S]. Jenis kedua, disebut [4Fe–4S], mengandung sebua kubus empat atom besi dan empat atom sulfur. Tiap-tiap atom pada gugus ini berkoordinasi dengan asam amino, biasanya koordinasi antara atom sulfur dengan sisteina. Kofaktor ion logam menjalani reaksi redoks tanpa mengikat ataupun melepaskan proton, sehingga pada rantai transpor elektron ia hanya berfungsi sebagai pengangkut elektron. Elektron bergerak cukup jauh melalui protein-protein ini dengan cara meloncat disekitar rantai kofaktor ini. Hal ini terjadi melalui penerowongan kuantum, yang terjadi dengan cepat pada jarak yang lebih kecil daripada 1,4×10−9 m.
Rantai transpor elektron eukariotik
Banyak proses katabolik biokimia, seperti glikolisis, siklus asam sitrat, dan oksidasi beta, menghasilkan koenzim NADH. Koenzim ini mengandung elektron yang memiliki potensial transfer yang tinggi. Dengan kata lain, ia akan melepaskan energi yang sangat besar semasa oksidasi. Namun, sel tidak akan melepaskan semua energi ini secara bersamaan karena akan menjadi reaksi yang tidak terkontrol. Sebaliknya, elektron dilepaskan dari NADH dan dipindahkan ke oksigen melalui serangkaian enzim yang akan melepaskan sejumlah kecil energi pada tiap-tiap enzim tersebut. Rangkaian enzim yang terdiri dari kompleks I sampai dengan kompleks IV ini disebut sebagai rantai transpor elektron dan ditemukan dalam membran dalam mitokondria. Suksinat juga dioksidasi oleh rantai transpor elektron, namun ia terlibat dalam lintasan yang berbeda.
Pada eukariota, enzim-enzim pada sistem transpor ini menggunakan energi yang dilepaskan dari oksidasi NADH untuk memompa proton melewati membran dalam mitokondria. Hal ini menyebabkan proton terakumulasi pada ruang antarmembran dan menghasilkan gradien elektrokimia di sepanjang membran. Energi yang tersimpan sebagai energi potensial ini kemudian digunakan oleh ATP sintase untuk menghasilkan ATP. Mitokondria terdapat pada hampir semua eukariota, dengan pengecualian pada protozoa anaerobik seperti Trichomonas vaginalis yang mereduksi proton menjadi hidrogen menggunakan hidrogenosom.
| Sistem pernapasen | Pasangan redoks | Potensial tengah
(Volt) |
|---|---|---|
| NADH dehidrogenase | NAD+ / NADH | −0.32 |
| Suksinat dehidrogenase | FMN atau FAD / FMNH2 atau FADH2 | −0.20 |
| Kompleks sitokrom bc1 | Koenzime Q10ox / Koenzime Q10red | +0.06 |
| Kompleks sitokrom bc1 | Sitokrom box / Sitokrom bred | +0.12 |
| Kompleks IV | Sitokrom cox / Sitokrom cred | +0.22 |
| Kompleks IV | Sitokrom aox / Sitokrom ared | +0.29 |
| Kompleks IV | O2 / HO- | +0.82 |
| Kondisi: pH = 7 | ||
Pada dasarnya, terdapat dua mekanisme katalitik yang dilakukan tiap kompleks enzim agar transfer elektron dapat menciptakan potensial membran, yaitu mekanisme iterasi redoks dan mekanisme pemompaan ion H+. Pada mekanisme iterasi redoks sendiri, reaksi reduksi akan mengikat ion H+, sedangkan reaksi oksidasi akan melepaskannya. Pada respirasi anaerobik, mekanisme yang sederhana ditunjukkan oleh format dehidrogenase dan nitrat reduktase yang terikat pada membran sel. Pada respirasi aerobik, mekanisme yang terjadi adalah sebagai berikut,
Kompleks I
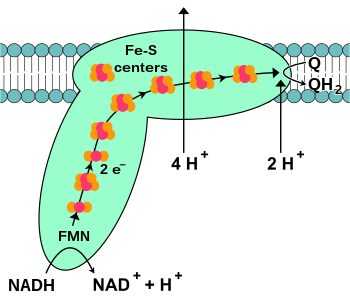
Kompleks I merupakan protein pertama pada rantai transpor elektron, berupa kompleks enzim yang disebut NADH-koenzim Q oksidoreduktase.
Pada hepatosit hewan sapi, kompleks I adalah enzim raksasa dengan 46 sub-unit dan massa molekul sekitar 1.000 kilodalton (kDa). Hanya struktur enzim kompleks I dari bakteri yang diketahui secara mendetail; pada kebanyakan organisme, kompleks ini menyerupai sepatu but dengan "bola" yang besar menyeruak keluar dari membran ke dalam mitokondria. Gen yang mengkode protein ini terdapat pada baik inti sel maupun genom mitokondria.
Reaksi redoks yang dikatalisis oleh enzim ini adalah oksidasi NADH, dan reduksi koenzim Q10 (diwakilkan dengan Q):

Oksidasi NADH akan menghasilkan NAD+ yang diperlukan untuk siklus asam sitrat dan oksidasi asam lemak,

Reaksi oksidasi NADH di atas dikopling oleh reaksi deiodinasi hormon tiroksin dengan promoter berupa peroksidase dan H2O2, sedangkan reduksi Q akan mentranspor elektron ke kompleks berikutnya hingga pada akhirnya digunakan untuk mereduksi oksigen menjadi air.

Awal mula reaksi terjadi ketika NADH berikatan dengan kompleks I dan menyumbang dua elektron. Elektron tersebut kemudian memasuki kompleks I via FMN, suatu gugus prostetik yang melekat pada kompleks. Tambahan elektron ke FMN mengubahnya menjadi bentuk tereduksi, FMNH2. Elektron kemudian ditransfer melalui rangkaian gugus besi-sulfur. Kemudian elektron ditransfer ke Q, mengubahnya menjadi QH2, dan menyebabkan 4 ion H+ terpompa keluar, menuju ke dalam sitoplasma, bukan ke dalam ruang antarmembran, oleh karena kompleks I terikat oleh 3 lapisan membran mitokondria. Pada sel prokariota Escherichia coli dan Klebsiella pneumoniae, kompleks I tidak meletupkan ion H+, melainkan ion Na+.
Terdapat baik jenis gugus besi-sulfur [2Fe-2S] maupun [4Fe–4S] dalam kompleks I.
Kopling yang terjadi dengan siklus asam sitrat,


Kompleks II

Kompleks II merupakan kompleks enzim yang disebut suksinat kuinon oksidoreduktase (EC 1.3.5.1) adalah titik masuk kedua pada rantai transpor elektron, yang terdiri dari kompleks enzim suksinat dehidrogenase aerobik dan fumarat reduktase anaerobik.
Kompleks II adalah satu-satunya kompleks enzim yang merupakan bagian dari kedua lintasan metabolisme, siklus asam sitrat maupun respirasi seluler pada rantai transpor elektron, dan terdiri dari empat subunit protein dan mengantung sebuah kofaktor flavin adenina dinukleotida yang terikat pada enzim, gugus besi-sulfur, dan sebuah gugus heme yang tidak berpartisipasi pada transfer elektron ke koenzim Q, namun dipercayai penting dalam penurunan produksi spesi oksigen reaktif. Enzim ini mereduksi fumarat menjadi suksinat dan meoksidasi hidrokuinon. Karena reaksi ini melepaskan energi lebih sedikit daripada oksidasi NADH, kompleks II tidak mentranspor proton melewati membran dan tidak berkontribusi terhadap gradien proton.
Reaksi redoks pada modus anaerobik oleh fumarat reduktase:
oksidasi

reduksi

Kopling yang terjadi dengan siklus asam sitrat,

Pada beberapa eukariota seperti cacing parasit Ascaris suum, terdapat enzim yang mirip dengan kompleks II, yaitu fumarat reduktase (menakuinol:fumarat oksidoreduktase, atau QFR). Kerja enzim ini terbalik dengan kerja kompleks II, yaitu mengoksidasi ubikuinol dan mereduksi fumarat. Hal ini mengizinkan cacing ini bertahan hidup dalam lingkungan anaerobik di usus besar dan menjalankan fosforilasi oksidatif anaerobik dengan fumarat sebagai akseptor elektron. Fungsi tak lazim kompleks II lainnya dapat dilihat pada parasit malaria Plasmodium falciparum. Pada organisme ini, fungsi kompleks II yang terbalik sebagai oksidase berperan penting dalam pemulihan ubikuinol, yang oleh parasit digunakan untuk biosintesis pirimidina.
Flavoprotein transfer elektron-Q oksidoreduktase
Pada kompleks II terdapat kompleks enzim ETF-QO dengan tiga domain pencerap, masing-masing mengikat FAD, kluster [4Fe-4S]1+, 2+ dan ubikuinon.
ETF-QO mempercepat reaksi redoks:
- reduksi senyawa Q-1 dengan elektron dari senyawa flavoprotein ET yang dapat berasal dari 11 macam flavoprotein dehidrogenase yang terdapat di dalam matriks mitokondria, Pada lintasan alternatif, elektron dapat mengalir dari kluster 4Fe4S dan dikatalitik oleh ETF-QO untuk mereduksi ubikuinon menjadi ubikuinol dengan koenzim FAD. Lintasan reaksi yang terjadi:
reduksi

oksidasi kofaktor

- oksidasi dengan substrat berupa asam lemak yang disebut lintasan oksidasi ß, katabolisme beberapa asam amino dan kolina, kemudian mentransfer elektronnya ke dalam kompleks II.
Pada mamalia, lintasan metabolisme ini sangat penting dan enzim yang berperan adalah asil-KoA dehidrogenase.
Reaksi yang terjadi:
oksidasi

reduksi kofaktor

Pada tumbuhan, ETF-QO juga penting dalam respon metabolik demi kelangsungan hidup tumbuhan pada periode lingkungan gelap yang berkepanjangan yang tidak memungkinkan terjadinya fotosintesis, sehingga terjadi simtoma hipoglisemia.
Kompleks III

Kompleks III juga dikenal sebagai kompleks enzim UCCR yang memiliki 11 berkas genetik UQCR. Pada mamalia, enzim ini berupa dimer, dengan tiap kompleks subunit mengandung 11 subunit protein, satu gugus besi-sulfur [2Fe-2S], dan tiga sitokrom yang terdiri dari satu sitokrom c1 dan dua sitokrom b. Sitokrom adalah sejenis protein pentransfer elektron yang mengandung paling tidak satu gugus heme. Atom besi dalam gugus heme kompleks III berubah dari bentuk tereduksi Fe (+2) menjadi bentuk teroksidasi Fe (+3) secara bergantian sewaktu elektron ditransfer melalui protein ini.
Reaksi yang dikatalisis oleh kompleks III adalah oksidasi satu molekul ubikuinol dan reduksi dua molekul sitokrom c. Tidak seperti koenzim Q yang membawa dua elektron, sitokrom c hanya membawa satu elektron.

Oleh karena hanya satu elektron yang dapat ditransfer dari donor QH2 ke akseptor sitokrom c, mekanisme reaksi kompleks III lebih rumit daripada kompleks lainnya, dan terjadi dalam dua langkah yang disebut siklus Q. Pada langkah pertama, enzim mengikat tiga substrat, pertama, QH2 yang akan dioksidasi kemudian dengan satu elektron dipindahkan ke sitokrom c yang merupakan substrat kedua. Dua proton yang dilepaskan dari QH2 dilepaskan ke dalam ruang antarmembran. Substrat ketiga adalah Q, yang menerima dua elektron dari QH2 dan direduksi menjadi Q.-, yang merupakan radikal bebas ubisemikuinon. Dua substrat pertama dilepaskan, namun zat antara ubisemikuinon ini tetap terikat. Pada langkah kedua, molekul kedua QH2 terikat dan kemudian melepaskan satu elektronnya ke akspetor sitokrom c. Elektron kedua dilepaskan ke ubisemikuinon yang terikat, mereduksinya menjadi QH2 ketika ia menerima dua proton dari matriks mitokondria. QH2 ini kemudian dilepaskan dari enzim.
Karena koenzim Q direduksi menjadi ubikuinol pada sisi dalam membran dan teroksidasi menjadi ubikuinon pada sisi luar, terjadi transfer proton di membran, yang menambah gradien proton. Mekanisme dua langkah ini sangat penting karena ia meningkatkan efisiensi transfer proton. Jika hanya satu molekul QH2 yang digunakan untuk secara langsung mereduksi dua molekul sitokrom c, efisiensinya akan menjadi setengah, dengan hanya satu proton yang ditransfer per sitokrom c yang direduksi.
Kompleks IV

Kompleks IV adalah protein terakhir pada rantai transpor elektron yang dikenal sebagai kompleks enzim COX.
Dari penelitian pada hepatosit hewan sapi, enzim ini memiliki struktur kompleks yang mengandung 13 subunit, antara lain 5 fosfatidil etanolamina, 3 fosfatidil gliserol, 2 asam kolat, 2 gugus heme A, dan beberapa kofaktor ion logam, meliputi tiga atom tembaga, satu atom magnesium, dan satu atom seng. Dua lintasan peletup ion H+ ditemukan membentang dari matriks hingga sitoplasma.
Pada model hepatosit hewan sapi, ion H+ dengan energi potensial elektrostatik berkisar antara 635meV, tampak dilepaskan dari sitokrom c oksidasefosfolipid vesikel (COV) pada kedua fase oksidatif dan reduktif, setelah dikirimkan dari proton loading site (PLS), pada saat ion H+ berikutnya tiba di PLS. Mekanisme yang ditunjukkan oleh peletupan ion H+ pada kompleks IV ini disebut efek Bohr redoks. Peletupan ion H+ (bahasa Inggris: deprotonation) terjadi bersamaan dengan perubahan gugus karboksil asam aspartat yang berada pada permukaan intermembran menjadi aspargina.
Enzim ini memediasi reaksi terakhir pada rantai transpor elektron dan mentransfer elektron ke oksigen, manakala memompa proton melewati membran. Oksigen yang menerima elektron, juga dikenal sebagai akseptor elektron terminal, direduksi menjadi air. Baik pemompaan proton secara langsung maupun konsumsi proton matriks pada reduksi oksigen berkontribusi kepada gradien proton. Menurut Keilin, reaksi yang dikatalisis oleh sitokrom c dan reduksi oksigennya adalah:

Reduktase dan oksidase alternatif
Enzim-enzim yang disebutkan di atas merupakan hasil kajian pada hewan mamalia. Sebenarnya, banyak organisme eukariotik lainnya yang memiliki rantai transpor elektron yang berbeda. Sebagai contoh, tumbuhan memiliki NADH oksidase alternatif, yang mengoksidasi NADH di sitosol daripada di matriks mitokondria, dan ia akan memindahkan elektron ke kolam ubikuinon. Enzim-enzim ini tidak mentranspor proton, sehingga ia mereduksi ubikuinon tanpa mengubah gradien elektronkimia membran dalam.
Contoh rantai transpor elektron divergen lainnya adalah oksidase alternatif yang ditemukan pada tumbuh-tumbuhan, beberapa spesies fungi, protista, dan kemungkinan pula pada beberapa hewan. Enzim ini secara langsung mentransfer elektron dari ubikuinol ke oksigen.
Lintasan tranpor elektron yang dihasilkan oleh NADH dan ubikuinon oksidase alternatif ini memiliki rendemen ATP yang lebih rendah. Keuntungan dari lintasan yang lebih singkat ini belumlah cukup jelas. Namun, oksidasi alternatif ini dihasilkan sebagai respon terhadap berbagai tekanan seperti hawa dingin, spesi oksigen reaktif, infeksi oleh patogen, dan faktor-faktor lainnya yang menghambat rantai transpor elektron secara penuh. Lintasan alternatif ini oleh karenanya akan meningkatkan resistansi organisme terhadap luka dengan menurunkan stres oksidatif.
Pengorganisasian kompleks-kompleks
Model awal bagaimana rantai kompleks respiratori ini terorganisasikan adalah bahwa kompleks-kompleks ini berdifusi dengan bebas dan terbebas dari membran mitokondria. Namun, data-data terbaru mensugestikan bahwa kompleks-kompleks ini kemungkinan membentuk struktur berorde tinggi yang disebut superkompleks ataupun "respirasom." Berdasarkan model superkompleks ini, berbagai jenis kompleks ini terdapat dalam bentuk sehimpunan enzim-enzim yang berinteraksi dan terorganisasi. Asosiasi ini mengizinkan penyaluran substrat di antara berbagai kompleks enzim, sehingga meningkatkan laju dan efisiensi transfer elektron. Dalam superkompleks mamalia, beberapa komponen kompleks akan lebih banyak daripada yang lainnya, dengan beberapa data mensugestikan rasio antara kompleks I/II/II/IV dan ATP sintase kira-kira 1:1:3:7:4. Walau demikian, perdebatan mengenai hipotesis superkompleks ini masihlah belum berakhir, karena beberapa data tampaknya tidak sesuai dengan model ini.
Rantai transpor elektron prokariotik
Berbeda dengan banyaknya kemiripan dalam struktur dan fungsi rantai transpor elektron pada eukariota, bakteri dan arkaea memiliki banyak jenis enzim transfer elektron yang sangat bervariasi. Enzim-enzim yang bervariasi ini pula menggunakan senyawa kimia yang bervaruasi sebagai substrat. Walau demikian, terdapat kesamaan dengan rantai transpor elektron eukarita, yaitu transpor elektron prokariotik juga menggunakan energi yang dilepaskan dari oksidasi substrat untuk memompa ion keluar masuk membran dan menghasilkan gradien elektrokimia. Fosforilasi oksidatif bakteri, utamanya bakteri Escherichia coli telah dipahami secara mendetail, manakala pada arkaea, hal ini masih belum dipahami dengan baik.
Perbedaan utama antara fosforilasi eukariotik dengan fosforilasi oksidatif prokariotik adalah bahwa bakteri dan arkaea menggunakan banyak senyawa-senyawa yang berbeda untuk menerima dan mendonor elektron. Hal ini sebenarnya mengizinkan prokariota untuk hidup dan tumbuh dalam berbagai jenis kondisi dan lingkungan. Pada E. coli, sebagai contohnya, fosforilasi oksidatif dapat didorong oleh sejumlah besar pasangan reduktor dan oksidator (lihat tabel di bawah). Potensial titik tengah suatu senyawa kimia mengukur seberapa banyak energi yang dilepaskan ketika ia dioksidasi maupun direduksi, dengan reduktor memiliki potensial negatif dan oksidator positif.
| Enzim pernapasen | Pasangan redoks | Potensial titik tengah
(Volt) |
|---|---|---|
| Format dehidrogenase | Bikarbonat / Format | −0,43 |
| Hidrogenase | Proton / Hidrogen | −0,42 |
| NADH dehidrogenase | NAD+ / NADH | −0,32 |
| Gliserol-3-fosfat dehidrogenase | DHAP / Gly-3-P | −0,19 |
| Piruvat oksidase | Asetat + Karbon dioksida / Piruvat | ? |
| Laktat dehidrogenase | Piruvat / Laktat | −0,19 |
| Asam D-amino dehidrogenase | 2-oksoasam + amonia / Asam D-amino | ? |
| Glukosa dehidrogenase | Glukonat / Glukosa | −0,14 |
| Suksinat dehidrogenase | Fumarat / Suksinat | +0,03 |
| Ubikuinol oksidase | Oksigen / Air | +0,82 |
| Nitrat reduktase | Nitrat / Nitrit | +0,42 |
| Nitrit reduktase | Nitrit / Amonia | +0,36 |
| Dimetil sulfoksida reduktase | DMSO / DMS | +0,16 |
| Trimetilamina N-oksida reduktase | TMAO / TMA | +0,13 |
| Fumarat reduktase | Fumarat / Suksinat | +0,03 |
Sebagaimana yang ditunjukkan oleh tabel di atas, E. coli dapat tumbuh dengan menggunakan reduktor seperti format, hidrogen, ataupun laktat sebagai donor elektron dan nitrat, DMSO, ataupun oksigen akseptor. Semakin besar perbedaan potensial titik tengah antra reduktor dan oksidator, semakin banyak pula energi yang dilepaskan ketika bereaksi. Dari seluruh pasangan senyawa ini, pasangan suksinat/fumarat tidak lazim karena potensial titik tengahnya mendekati nol. Suksinat oleh karenanya dapat dkoksidasi menjadi fumarat apabila terdapat oksidator kuat seperti oksigen dan fumarat dapat direduksi menjadi suksinat menggunakan reduktor kuat seperti format. Reaksi alternatif ini dikatalisis oleh suksinat dehidrogenase untuk oksidasi suksinat dan fumarat reduktase untuk reduksi fumarat.
Beberapa prokariota menggunakan pasangan redoks yang hanya memiliki perbedaan potensial titik tengah yang kecil. Sebagai contohnya, bakteri yang melakukan nitrifikasi seperti Nitrobakter mengoksidasi nitrit menjadi nitrat dan mendonarkan elektron ke oksigen. Sejumlah kecil energi yang dilepaskan oleh reaksi ini cukup untuk memompa proton dan menghasilkan ATP, namun tidak cukup untuk menghasilkan NADH ataupun NADPH secara langsung untuk digunakan dalam anabolisme. Permasalahan ini diselesaikan dengan menggunakan nitrit oksidoreduktase untuk menghasilkan gaya gerak proton yang cukup untuk menjalankan sebagai rantai transpor elektron secara terbalik, menyebabkan kompleks I memproduksi NADH.
Prokariota mengontrol penggunaan donor dan akseptor elektron ini dengan memproduksi enzim tertentu sesuai dengan kondisi lingkungan. Fleksibilitas ini dimungkinkan karena oksidase dan reduktase yang berbeda menggunakan kolam ubikuinon yang sama. Ini mengizinkan banyak kombinasi enzim untuk bekerja secara bersamaan, yang saling terhubung oleh zat antara ubikuinol.
Selain beranekaragamnya lintasan metabolisme ini, prokariota juga memiliki sejumlah besar isozim, yaitu enzim-enzim berbeda yang mengkatalisis reaksi yang sama. Sebagai contohnya, E. coli memiliki dua jenis ubikuinol oksidase yang berbeda. Di bawah kondisi aerob, sel menggunakan oksidase yang berafinitas rendah terhadap oksigen yang dapat mentranspor dua proton per elektron. Namun, apabila kadar oksigen menurun, sel akan menggunakan oksidase yang hanya mentransfer satu proton per elektron namun berafinitas tinggi terhadap oksigen.
ATP sintase (kompleks V)
ATP sintase, juga disebut kompleks V, adalah enzim terakhir dalam lintasan fosforilasi oksidatif. Enzim ini ditemukan di seluruh organisme hidup dan berfungsi sama pada prokariota maupun eukariota. Enzim ini menggunakan energi yang tersimpan pada gradien proton di sepanjang membran untuk mendorong sintesis ATP dari ADP dan fosfat (Pi). Perkiraan jumlah proton yang diperlukan untuk mensintesis satu ATP berkisar antara tiga sampai dengan empat, dengan beberapa peneliti yang mensugestikan bahwa sel dapat memvariasikan rasio ini sesuai dengan kondisi.

Reaksi fosforilasi ini adalah reaksi kesetimbangan, yakni ia dapat digeser dengan mengubah gaya gerak proton. Dengan ketiadaan gaya gerak proton, reaksi ATP sintase akan berjalan dari sisi kanan ke kiri, menghidrolisis ATP dan memompa proton keluar dari matriks melewati membran. Namun, ketika gaya gerak protonnya tinggi, reaks dipaksa untuk berjalan secara terbalik, yaitu dari sisi kanan ke kiri, mengizinkan proton mengalir dan mengubah ADP menjadi ATP.
ATP sintase adalah sebuah kompleks protein yang besar dengan bentuk seperti jamur. Kompleks enzim ini pada mamalia mengandung 16 subunit dan memiliki massa kira-kira 600 kilodalton. Bagian yang tertanam pada membran disebut FO dan mengandung sebuah cincin subunit c dan saluran proton. "Tangkai" dan kepala yang berbentuk bola disebut F1 dan merupakan tempat sintesis ATP. Kompleks yang berbentuk bola pada ujung akhir F1 mengandung enam protein yang dapat dibagi menjadi dua jenis: tiga subunit α dan tiga subunit β), manakala bagian "tangkai" terdiri dari satu protein: subunit γ, dengan ujung tangkai menusuk ke dalam bola subunit α dan β. Baik subunit α dan β mengikat nukleotida, namun hanya subunit β yang mengkatalisis reaksi sintesis ATP. Di samping F1 pula terdapat sebuah subunit berbentuk batang yang menghubungakan subunit α dan β dengan dasar enzim.
Seiring dengan mengalirnya proton melewati membran melalui saluran ini, motor FO berotasi. Rotasi dapat disebabkan oleh perubahan pada ionisasi asam amino cincin subunit c, menyebabkan interaksi elektrosatik yang menolak cincin subunit c. Cincin yang berotasi ini pada akhirnya akan memutar "as roda" (tangkai subunit γ). Subunit α dan β dihalangi untuk berputar oleh batang samping yang berfungsi sebagai stator. Pergerakan ujung subunit γ yang berada dalam bola subunit α dan β memberikan energi agar tapak aktif pada subunit β menjalankan siklus pergerakan yang memproduksi dan kemudian melepaskan ATP.

Reaksi sintesis ATP ini disebut sebagai mekanisme perubahan ikatan (binding change mechanism) dan melibatkan tapak aktif subunit β yang berputar terus dalam tiga keadaan. Pada keadaan "terbuka", ADP dan fosfat memasuki tapa aktif (ditunjukkan dalam warna coklat pada diagram). Protein kemudian menutup dan mengikat ADP dan fosfat secara longgar (keadaan "longgar" ditunjukkan dalam warna merah). Enzim kemudian berubah bentuk lagi dan memaksa kedua molekul ini bersama, dengan tapak aktif dalam keadaan "ketat" (ditunjukan dalam warna merah jambu) dan mengikat molekul ATP yang terbentuk. Tapak aktif kemudian kembali lagi ke keadaan terbuka dan melepaskan ATP untuk kemudian mengikat ADP dan fosfat, dan memulai siklus yang baru.
Pada beberapa bakteri dan arkaea, sintesis ATP didorong oleh pergerakan ion natrium yang melalui membran sel daripada pergerakan proton. Arkaea seperti Methanococcus juga mengandung A1Ao sintase, sebuah bentuk enzim yang mengandung protein tambahan dengan kemiripan urutan asam amino yang kecil dengan subunit ATP sintase bakteri dan eukariota lainnya. Adalah mungkin bahwa pada beberapa spesies, bentuk enzim A1Ao adalah ATP-sintase terspesialisasi yang digerakkan oleh natrium, namun ini tidaklah benar pada keseluruhan kasus.
Spesi oksigen reaktif
Oksigen molekuler merupakan akseptor elektron akhir yang ideal, karena ia merupakan oksidator kuat. Reduksi oksigen melibatkan zat antara yang berpotensi bahaya. Walaupun transfer empat elektron dan empat proton akan mereduksi oksigen menjadi air, yang tidak berbahaya, transfer satu atau dua elektron akan menghasilkan anion superoksida ataupun anion peroksida, yang sangat reaktif dan berbahaya.

Spesi oksigen reaktif dan produk reaksinya ini seperti radikal hidroksil, sangatlah berbahaya bagi sel, karena akan mengoksidasi protein dan mengakibatkan mutasi pada DNA. Kerusakan ini berkontribusi terhadap penyakit dan diajukan pula merupakan salah satu akibat dari penuaan.
Kompleks sitokrom c sangat efisien mereduksi oksigen menjadi air, dan melepaskan hanya sedikit zat antara yang tereduksi secara parsial. Namun terdapat sejumlah kecil anion superoksida dan peroksida yang diproduksi oleh rantai transpor elektron. Terutama pentingnya adalah pada reduksi koenzime Q pada kompleks III, karena radikal bebas ubikuinon yang sangat reaktif terbentuk sebagai zat antara dalam siklus Q. Spesi yang tidak stabil ini dapat menyebabkan "kebocoran" elektron ketika elektron ditransfer secara langsung ke oksigen dan menghasilkan superoksida. Karena laju produksi spesi oskigen reaktif oleh kompleks pemompa proton ini tertinggi ketika potensial membran tinggi, diajukan bahwa mitokondria meregulasi aktivitas kompleks untuk menjaga potensial membran berada dalam kisaran yang kecil sehingga menyeimbangkan produksi ATP terhadap produksi oksidator. Sebagai contohnya, oksidator dapat mengaktivasi UCP (uncoupling protein) yang menurunkan potensial membran.
Untuk melawan spesi oksigen reaktif ini, sel mengandung sejumlah sistem antioksidan, meliputi vitamin antioksidan seperti vitamin C dan vitamin E, dan enzim antioksidan seperti superoksida dismutase, katalase, dan peroksidase, yang menetralkan spesi reaktif sehingga mengurangi kerusakan sel.
Gugus anion superoksida, senyawa organik yang sangat aktif yang terdapat pada molekul hidrogen peroksida dan jenis ROS lainnya, merupakan produk samping reaksi redoks yang terjadi pada rantai transpor elektron. Gangguan mekanisme produksi ROS dapat berakibat pada berbagai macam patologi seperti diabetes, neurodegenerasi, gagal jantung, chronic obstructive pulmonary disease. Domain produksi anion superoksida terletak pada kompleks I dan kompleks III.
Dari beberapa senyawa intermediat pengusung satu elektron, radikal bebas SQ- dianggap merupakan senyawa yang paling berperan aktif dalam mereduksi molekul oksigen menjadi anion superoksida. Molekul semikuinon dihasilkan kompleks I dan III sebagai hasil reduksi ubikuinon atau oksidasi ubikuinol,

SQ- akan melekat pada kompleks I atau III hingga saat terstimulasi elektron yang kedua dengan reaksi,

Semikuinon lebih lanjut dapat berinteraksi langsung dengan molekul oksigen dengan reaksi,

Inhibitor
Terdapat beberapa obat dan racun yang dikenal baik menginhibisi fosforilasi oksidatif. Walaupun semua racun hanya menginhibisi satu enzim pada rantai transpor elektron, inhibisi pada langkah apapun pada proses ini akan menghentikan keseluruhan proses. Contohnya, jika oligomisin menginhibisi ATP sintase, proton tidak dapat mengalir balik ke dalam mitokondria. Akibatnya, pompa proton tidak dapat bekerja, karena gradien konsentrasinya menjadi terlalu kuat untuk diatasi. NADH kemudian tidak akan lagi teroksidasi dan siklus asam sitrat berhenti bekerja karena konsentrasi NAD+ menurun di bahwa kadar yang cukup agar enzim bekerja.
| Senyawa | Kegunaan | Efek terhadap fosforilasi oksidatif |
|---|---|---|
|
Sianida Karbon monoksida |
Racun | Menghambat rantai transpor elektron dengan terikat lebih kuat daripada oksigen pada pusat Fe–Cu dalam sitokrom c oksidase, mencegah reduksi oksigen. |
| Oligomisin | Antibiotik | Menghambat ATP sintase dengan memblokir aliran proton ke subunit Fo. |
|
CCCP 2,4-Dinitrofenol |
Racun | Ionofor yang mengganggu gradien proton dengan membawa proton melewati membran. Ionofor ini mengawagandengkan (uncouple) pompa proton dari sintesis ATP karena ia membawa proton melewati membran mitokondria dalam. |
| Rotenon | Pestisida | Mencegah transfer elektron dari kompleks I ke ubikuinon dengan menutup tapak ikat ubikuinon. |
| Malonat dan oksaloasetat | Inhibitor kompetitif suksinat dehidrogenase (kompleks II). |
Tidak semua inhibitor fosforilasi oksidatif bersifat racun. Pada jaringan lemak coklat, saluran proton yang diregulasi disebut UCP (uncoupling protein), yang dapat mengawagandengkan respirasi dari sintesis ATP. Respirasi cepat ini menghasilkan panas, dan proses ini sangat penting dalam menjaga suhu tubuh pada hewan yang berhibernasi, walaupun protein ini kemungkinan juga memiliki fungsi umum dalam respon sel terhadap stres.
Bacaan lebih lanjut
Pengenalan
- Nelson DL (2004). Lehninger Principles of Biochemistry (edisi ke-4th). W. H. Freeman. ISBN 0-716-74339-6.
- Schneider ED (2006). Into the Cool: Energy Flow, Thermodynamics and Life (edisi ke-1st). University of Chicago Press. ISBN 0-226-73937-6.
- Lane N (2006). Power, Sex, Suicide: Mitochondria and the Meaning of Life (edisi ke-1st). Oxford University Press, USA. ISBN 0199205647.
Lanjutan
- Nicholls DG (2002). Bioenergetics 3 (edisi ke-1st). Academic Press. ISBN 0-125-18121-3.
- Haynie D (2001). Biological Thermodynamics (edisi ke-1st). Cambridge University Press. ISBN 0-521-79549-4.
- Rajan SS (2003). Introduction to Bioenergetics (edisi ke-1st). Anmol. ISBN 8-126-11364-2.
- Wikstrom M (Ed) (2005). Biophysical and Structural Aspects of Bioenergetics (edisi ke-1st). Royal Society of Chemistry. ISBN 0-854-04346-2.
Pranala luar
Informasi umum
- Animated diagrams illustrating oxidative phosphorylation Wiley and Co Concepts in Biochemistry
- Oxidative phosphorylation Diarsipkan 1999-10-12 di Wayback Machine. Metabolic Pathways of Biochemistry, the online reference of metabolism for students and teachers, at George Washington University
- ATP synthase - the rotary engine in the cell Diarsipkan 2010-10-25 di Wayback Machine. Brief introduction, including videos of microscope images of the enzyme rotating, at Tokyo Institute of Technology
- On-line biophysics lectures Antony Crofts, University of Illinois at Urbana-Champaign
Informasi struktural
- Animations of the ATP synthase Diarsipkan 2009-01-20 di Wayback Machine. Hongyun Wang and George Oster, University of California, Berkeley
-
PDB molecule of the month:
- ATP synthase Diarsipkan 2020-07-24 di Wayback Machine.
- Cytochrome c Diarsipkan 2020-07-24 di Wayback Machine.
- Cytochrome c oxidase Diarsipkan 2020-07-24 di Wayback Machine.
- Interactive molecular models at Universidade Fernando Pessoa:
- NADH dehydrogenase Diarsipkan 2009-01-12 di Wayback Machine.
- succinate dehydrogenase Diarsipkan 2009-01-11 di Wayback Machine.
- Coenzyme Q - cytochrome c reductase Diarsipkan 2009-01-12 di Wayback Machine.
- cytochrome c oxidase Diarsipkan 2009-01-11 di Wayback Machine.