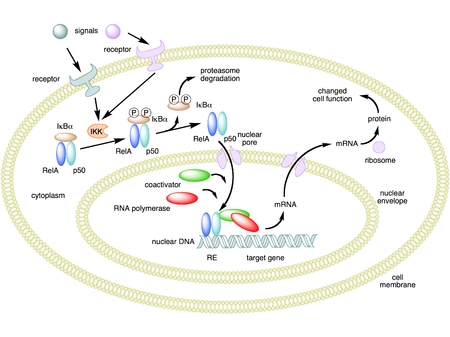
Mekanisme aksi NF-κB. Pada gambar ini, heterodimer NF-κB antara protein Rel dan p50 digunakan sebagai contoh. Saat keadaan tidak aktif, NF-κB berada di sitosol yang membentuk kompleks dengan protein penghambat
IκBα. Melalui intermediasi reseptor membran integral, berbagai sinyal ekstraselular dapat mengaktifkan enzim
IκB kinase (IKK). IKK, pada gilirannya, memfosforilasi protein IκBα, yang menghasilkan
ubiquitinasi, disosiasi IκBα dari NF-κB, dan akhirnya degradasi IκBα oleh
proteosom. NF-κB yang diaktifkan kemudian pindah ke dalam nukleus tempat ia mengikat urutan spesifik DNA yang disebut respon elemen (RE). Kompleks DNA/NF-κB kemudian merekrut protein lain seperti
koaktivator dan
RNA polimerase, yang mentranskripsi DNA ke dalam mRNA. Selanjutnya, mRNA diterjemahkan ke dalam protein, menghasilkan perubahan fungsi sel.

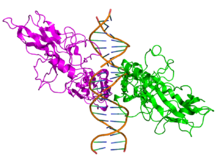
Diagram skematika struktur protein NF-κB. Ada dua kelas struktural protein NF-κB: kelas I (atas) dan kelas II (bawah). Kedua kelas protein mengandung N-terminal
domain pengikat DNA (DBD), yang juga berfungsi sebagai antarmuka dimerisasi ke faktor transkripsi NF-κB lainnya dan juga mengikat protein penghambat
IκBα. C-terminal protein kelas I berisi sejumlah
pengulangan ankyrin dan memiliki aktivitas transrepresi. Sebaliknya, protein C kelas II memiliki fungsi
transaktivasi.
NF-κB (bahasa Inggris: nuclear factor kappa-light-chain-enhancer of activated B cells, nuclear factor-kappaB) adalah keluarga faktor transkripsi yang berperan dalam respon seluler terhadap rangsangan seperti stres, sitokina, radikal bebas, logam berat, iradiasi ultraviolet, LDL yang teroksidasi, dan bakteri atau virus (antigen). NF-κB memainkan peran kunci dalam mengatur respon imun terhadap infeksi (rantai ringan κ adalah komponen penting imunoglobulin). Regulasi NF-κB yang salah telah dikaitkan dengan kanker, inflamasi dan penyakit autoimun, syok septik, infeksi virus, dan gangguan sistem imun. NF-κB juga terlibat dalam proses plastisitas sinaptik dan memori.
Penemuan
NF-κB ditemukan oleh Dr. Ranjan Sen di laboratorium penerima Hadiah Nobel David Baltimore melalui interaksinya dengan urutan pasangan 11 basa pada enhancer rantai ringan imunoglobulin pada sel B.
Sejak ditemukan pada tahun 1986, banyak studi menunjukkan hubungan antara lintasan sinyal NF-κB dan respon peradangan:
Struktur
Semua protein keluarga NF-κB berbagi domain homologi Rel di bagian N-terminal. Sebuah subfamili protein NF-κB, dengan anggota RelA, RelB, dan c-Rel, memiliki domain transaktivasi di bagian C-terminal. Sebaliknya, protein NF-κB1 dan NF-κB2 disintesis sebagai prekursor besar, p105 dan p100, yang menjalani proses untuk menghasilkan subunit NF-κB matang, p50 dan p52. Pengolahanam p105 dan p100 diperantarai oleh jalur ubiquitin / proteasom dan melibatkan degradasi selektif daerah C-terminal yang mengandung pengulangan ankyrin. Sedangkan pembuatan p52 dari p100 adalah proses yang diatur ketat, dan p50 dihasilkan dari pemrosesan konstitutif p105. Protein p50 dan p52 tidak memiliki kemampuan intrinsik untuk mengaktifkan transkripsi dan oleh karena itu telah diusulkan untuk beraksi sebagai represor transkripsional saat mengikat elemen κB sebagai homodimer. Memang, hal ini membingungkan pada interpretasi penelitian p105-knockout, yang mana manipulasi genetik menyingkirkan IκB (p105 penuh) dan kemungkinan represor (p50 homodimer) selain aktivator transkripsi (heterodimer RelA-p50).
Anggota
Anggota keluarga NF-κB berbagi struktur homologi dengan v-Rel onkoprotein retroviral, yang menghasilkan klasifikasi sebagai protein NF-kB/Rel.
Ada lima protein dalam famili NF-κB mamalia:
| Kelas
|
Protein
|
Alias
|
Gen
|
| I
|
NF-κB1
|
p105 → p50
|
NFKB1
|
| NF-κB2
|
p100 → p52
|
NFKB2
|
| II
|
RelA
|
p65
|
RELA
|
| RelB
|
|
RELB
|
| c-Rel
|
|
REL
|
Berikut adalah lima anggota keluarga NF-κB manusia (gambar ada di samping kanan):
Pensinyalan
Aktivasi NF-κB terjadi melalui dia jalur utama: jalur kanonikal dan non-kanonikal. Jalur kanonikal memperantarai aktivasi NF-κB1 p50, RELA dan c-REL (yang disebut juga anggota keluarga NF-κB kanonikal), sedangkan jalur non-kanonikal secara selektif mengaktifkan anggota NF-κB terikat p100, utamanya NF-κBB2 p52 dan RELB (biasa disebut anggota keluarga NF-κB non-kanonikal).
Proses aktivasi (kanonikal/klasikal)
Aktivasi NF-κB diinisiasi oleh degradasi protein IκB dalam merespon suatu stimulus sinyal. Hal ini terjadi terutama melalui aktivasi kinase yang disebut IκB kinase (IKK). IKK terdiri dari heterodimer dari subunit IKKα dan IKKβ katalitik dan protein pengatur "utama" yang disebut NEMO (modulator penting NF-κB) atau gamma IKK. Bila diaktifkan oleh sinyal, biasanya berasal dari luar sel, kinase IκB memfosforilasi dua residu serin yang terletak di domain pengaturan IκB. Ketika fosforilasi pada serin ini (misalnya, serine 32 dan 36 pada manusia IκBα), protein IκB dimodifikasi oleh proses yang disebut ubikuitinasi, yang kemudian menyebabkannya terdegradasi oleh organel sel yang disebut proteasom.
Non-kanonikal/jalur alternatif
Pengaktifan jalur NF-κB non-kanonikal didasarkan pada pemrosesan p100. Berbeda dengan pemrosesan konstitutif p105, pemrosesan p100 dikontrol dengan ketat dan terjadi dengan cara diinduksi. Pengolahan p100 yang diinduksi sinyal mengarah pada pembentukan NF-κB2 p52 serta translokasi inti anggota NF-κB p100, yang sebagian besar adalah RELB dan NF-κB2 p52. Pemrosesan p100 diinduksi melalui fosforilasi pada residu serin C-terminal spesifik (serine 866 dan 870), yang memicu ubiluitilasi p100 melalui perekrutan E3 ubiquitin ligase βTrCP. Komponen pensinyalan sentral dari jalur NF-κB non-kanonikal adalah NF-κB-inducing kinase (NIK; juga dikenal sebagai MAP3K14), yang menginduksi fosforilasi p100 melalui aktivasi kinase IKKα1. Berbeda dengan aktivasi jalur NF-κB kanonikal yang cepat dan sementara, aktivasi jalur NF-κB non-kanonikal sangat lambat dan terus-menerus. Pendorong khas jalur NF-κB non-kanonikal adalah ligan dari subset anggota superfamili reseptor faktor nekrosis tumor (TNFR).
|
|---|
| Perpustakaan nasional |
|
| Lain-lain |
|