

Evolusi manusia adalah proses evolusi dalam sejarah primata yang menyebabkan munculnya Homo sapiens sebagai spesies tersendiri dari famili hominid, yang mencakup kera besar. Proses ini meliputi perkembangan bertahap dari sifat-sifat seperti bipedalisme dan kemampuan berbahasanya manusia, serta perkawinan silang dengan hominin lain, yang menunjukkan bahwa evolusi manusia tidaklah linear, akan tetapi berbentuk seperti jaringan. Studi evolusi manusia melibatkan sejumlah disiplin ilmu, termasuk antropologi fisik, antropologi evolusi, primatologi, arkeologi, paleontologi, neurobiologi, etologi, linguistik, psikologi evolusioner, embriologi dan genetika.
Primata berpisah dari mamalia lain sekitar 85 juta tahun silam, yakni pada periode Kapur Akhir. Primata kemudian menghasilkan garis keturunan yang berurutan mengarah ke superfamili kera, yang memunculkan keluarga hominid dan ungka; yang kemudian berpisah satu sama lain sekitar 15–20 juta tahun yang lalu. Hominid Asia (mencakup orang utan) lalu berpisah dari hominid Afrika sekitar 14 juta tahun silam. Dari hominid Afrika, Hominin (termasuk sub-tribus Australopithecine dan Pan) berpisah dari tribus Gorillini (gorila) antara 8–9 juta tahun silam; Australopithecine (termasuk nenek moyang berkaki dua manusia yang telah punah) berpisah dari genus Pan (yang terdiri dari simpanse dan bonobo) 4–7 juta tahun silam. Dari hasil evolusi Australopithecine kemudian lahirlah genus Homo dengan ditandai oleh kemunculan H. habilis lebih dari 2 juta tahun silam, dan dari garis keturunan tersebut kemudian lahirlah manusia modern secara anatomis sekitar 300.000 tahun silam.
Sebelum Homo
Evolusi awal Primata
Sejarah evolusi primata dapat ditelusuri kembali 65 juta tahun yang lalu. Salah satu spesies mamalia mirip primata tertua yang diketahui, Plesiadapis, berasal dari Amerika Utara; yang lainnya, Archicebus, berasal dari China. Primata basal serupa lainnya tersebar luas di Eurasia dan Afrika selama kondisi tropis Paleosen dan Eosen.

David R. Begun menyimpulkan bahwa primata purba berkembang di Eurasia dan bahwa garis keturunan yang mengarah ke kera dan manusia Afrika, termasuk Dryopithecus, bermigrasi ke selatan dari Eropa atau Asia Barat ke Afrika. Populasi primata tropis yang masih ada—yang terlihat paling lengkap di lapisan fosil Eosen Atas dan Oligosen paling bawah dari depresi Faiyum di barat daya Kairo—memunculkan semua spesies primata yang masih ada, termasuk lemur di Madagaskar, kukang di Asia Tenggara, galago atau "bayi semak" di Afrika, dan anthropoid, yaitu Platyrrhines atau monyet Dunia Baru, Catarrhines atau monyet Dunia Lama, dan kera besar, termasuk manusia dan hominid lainnya.
Catarrhini paling purba yang diketahui adalah Kamoyapithecus dari Oligosen paling atas di Eragaleit di Lembah Celah Besar utara di Kenya, yang berasal dari 24 juta tahun yang lalu. Nenek moyangnya diperkirakan merupakan spesies yang terkait dengan Aegyptopithecus, Propliopithecus, dan Parapithecus dari Faiyum, sekitar 35 juta tahun yang lalu. Pada tahun 2010, Saadanius dideskripsikan sebagai kerabat dekat dari nenek moyang bersama terakhir dari kelompok mahkota catarrhines, dan secara tentatif bertanggal 29-28 juta tahun yang lalu, membantu mengisi celah 11 juta tahun dalam catatan fosil.

Pada Miosen Awal, sekitar 22 juta tahun yang lalu, banyak jenis catarrhini primitif dari Afrika Timur yang beradaptasi secara arboreal. Hal ini menunjukkan sejarah panjang diversifikasi awal. Fosil-fosil pada 20 juta tahun yang lalu meliputi fragmen-fragmen yang berkaitan dengan Victoriapithecus, monyet Dunia Lama yang paling awal. Beberapa genera yang diperkirakan masuk dalam garis keturunan kera yang berasal lebih dari 13 juta tahun yang lalu adalah Proconsul, Rangwapithecus, Dendropithecus, Limnopithecus, Nacholapithecus, Equatorius, Nyanzapithecus, Afropithecus, Heliopithecus, dan Kenyapithecus, semuanya berasal dari Afrika Timur.
Kehadiran non-cercopithecid umum lainnya dari Miosen Tengah dari situs-situs yang jauh—Otavipithecus dari endapan gua di Namibia, dan Pierolapithecus dan Dryopithecus dari Perancis, Spanyol dan Austria—adalah bukti dari keanekaragaman bentuk yang luas di seluruh Afrika dan cekungan Mediterania selama rezim iklim yang relatif hangat dan seimbang pada Miosen Awal dan Tengah. Hominoid Miosen termuda, Oreopithecus, berasal dari lapisan batu bara di Italia yang berasal dari 9 juta tahun yang lalu.
Bukti molekuler mengindikasikan bahwa garis keturunan owa (famili Hylobatidae) berpisah dari garis keturunan kera besar sekitar 18-12 juta tahun lalu, dan garis keturunan orangutan (subfamili Ponginae) berpisah dari kera besar lainnya sekitar 12 juta tahun; Tidak ada fosil yang secara jelas mendokumentasikan nenek moyang owa, yang mungkin berasal dari populasi hominoid Asia Tenggara yang sejauh ini belum diketahui, tetapi fosil proto-orangutan mungkin diwakili oleh Sivapithecus dari India dan Griphopithecus dari Turki, yang berasal dari sekitar 10 juta tahun yang lalu.
Homininae (hominid Afrika) yang merupakan subfamili dari Hominidae berpisah dari Ponginae (orang utan) sekitar 14 juta tahun yang lalu. Hominin (termasuk manusia dan sub-suku Australopithecine dan Panina) berpisah dari suku Gorillini (gorila) antara 8-9 juta tahun lalu; Australopithecine (termasuk nenek moyang berkaki dua yang telah punah) berpisah dari genus Pan (yang terdiri dari simpanse dan bonobo) 4-7 juta tahun lalu. Dari Australopithecine yang berevolusi kemudian muncullah genus Homo yang ditandai dengan lahirnya spesies H. habilis lebih dari 2 juta tahun yang lalu, sementara manusia modern secara anatomis kemudian lahir dari garis keturunan tersebut di Afrika sekitar 300.000 tahun yang lalu.
Perpisahan klade manusia dari kera besar lainnya
Spesies yang dekat dengan nenek moyang bersama terakhir gorila, simpanse, dan manusia mungkin diwakili oleh fosil Nakalipithecus yang ditemukan di Kenya dan Ouranopithecus yang ditemukan di Yunani. Bukti molekuler menunjukkan bahwa antara 8 dan 4 juta tahun yang lalu, pertama gorila, dan kemudian simpanse (genus Pan) memisahkan diri dari garis yang mengarah ke manusia. DNA manusia sekitar 98,4% identik dengan simpanse ketika membandingkan polimorfisme nukleotida tunggal (lihat genetika evolusi manusia). Namun, catatan fosil gorila dan simpanse terbatas; pengawetan fosil keduanya buruk, diakibatkan tanah hutan hujan yang cenderung asam yang mana melarutkan tulang belulang, dan bias penyampelan kemungkinan turut berkontribusi pada masalah ini.
Hominin-hominin lain mungkin beradaptasi dengan lingkungan yang lebih kering di luar sabuk khatulistiwa; dan di sana mereka bertemu antelop, hyena, anjing, babi, gajah, kuda, dan hewan-hewan lainnya. Sabuk khatulistiwa menyusut setelah sekitar 8 juta tahun lalu, dan hanya ada sedikit bukti fosil untuk divergensi garis keturunan hominin dari garis keturunan gorila dan simpanse yang diperkirakan terjadi sekitar waktu itu. Fosil paling awal yang dianggap oleh beberapa ahli sebagai bagian dari garis keturunan manusia adalah Sahelanthropus tchadensis (7 Ma) dan Orrorin tugenensis (6 Ma), diikuti oleh Ardipithecus (5,5-4,4 Ma), dengan spesies Ar. kadabba dan Ar. ramidus.
Dalam studi sejarah kehidupan Ar. ramidus, telah disebutkan bahwa spesies ini memberikan bukti adanya serangkaian adaptasi anatomi dan perilaku pada hominin purba yang tidak ditemukan pada spesies kera besar lain yang masih ada. Studi ini menunjukkan adanya kedekatan antara morfologi tengkorak Ar. ramidus dengan bayi dan simpanse remaja, yang menunjukkan bahwa spesies ini mengembangkan morfologi kraniofasial remaja atau paedomorfik melalui disosiasi heterokronik dari jalur pertumbuhan. Juga dikatakan bahwa spesies ini mendukung gagasan bahwa hominin yang sangat awal, mirip dengan bonobo (Pan paniscus) spesies yang kurang agresif dari genus Pan, mungkin telah berevolusi melalui proses domestikasi diri. Oleh karena itu, dengan argumen menentang apa yang disebut "model referensial simpanse" para penggagas berpendapat bahwa penggunaan perilaku sosial dan perkawinan simpanse (Pan troglodytes) tidak dapat lagi digunakan dalam model evolusi sosial hominin purba. Ketika mengomentari ketiadaan morfologi taring agresif pada Ar. ramidus dan implikasinya terhadap evolusi psikologi sosial hominin, mereka menulis:
Tentu saja Ar. ramidus berbeda secara signifikan dari bonobo, bonobo telah mempertahankan kompleks pengasahan gigi taring fungsional. Namun, fakta bahwa Ar. ramidus memiliki kesamaan dengan bonobo dalam penurunan dimorfisme seksual, dan bentuk yang lebih paedomorfik dibandingkan dengan simpanse, menunjukkan bahwa adaptasi perkembangan dan sosial yang terbukti ada pada bonobo mungkin dapat membantu rekonstruksi psikologi sosial dan seksual pada hominin awal di masa depan. Bahkan trend ke arah meningkatnya perawatan oleh ibu, pemilihan pasangan oleh betina, dan domestikasi diri mungkin lebih kuat dan lebih halus pada Ar. ramidus daripada apa yang kita lihat pada bonobo.
Para penyusun penelitian ini berpendapat bahwa banyak adaptasi dasar manusia berkembang di ekosistem hutan kuno di akhir Miosen dan awal Pliosen Afrika. Alhasil, mereka berpendapat bahwa manusia mungkin tidak mewakili evolusi dari nenek moyang mirip simpanse seperti yang selama ini diduga. Hal ini menunjukkan bahwa banyak adaptasi manusia modern mewakili sifat-sifat yang mendalam secara filogenetik dan bahwa perilaku dan morfologi simpanse mungkin berkembang setelah mereka berpisah dari nenek moyang bersama mereka dengan manusia.
Genus Australopithecus

Genus Australopithecus berevolusi di Afrika timur sekitar 4 juta tahun yang lalu sebelum menyebar ke seluruh Afrika dan akhirnya punah 2 juta tahun yang lalu. Selama periode waktu ini terdapat berbagai bentuk Australopithecus, termasuk Australopithecus anamensis, Au. afarensis, Au. sediba, dan Au. africanus. Masih terdapat beberapa perdebatan di kalangan akademisi apakah spesies hominid Afrika tertentu pada masa ini, seperti Au. robustus dan Au. boisei, merupakan anggota dari genus yang sama; jika demikian, mereka dianggap tergolong Au. robust australopith sementara yang lainnya dianggap sebagai Au. glacile australopith. Namun, jika spesies-spesies ini rupanya bagian dari genus tersendiri, maka mereka dapat diberi nama tersendiri, seperti Paranthropus.
- Australopithecus (4–1.8 Ma), dengan spesies Au. anamensis, Au. afarensis, Au. africanus, Au. bahrelghazali, Au. garhi, and Au. sediba;
- Kenyanthropus (3–2.7 Ma), dengan spesies K. platyops;
- Paranthropus (3–1.2 Ma), dengan spesies P. aethiopicus, P. boisei, and P. robustus
Spesies baru yang diajukan, Australopithecus deyiremeda dianggap hidup pada periode waktu yang sama dengan Au. afarensis. Ada perdebatan apakah Au. deyiremeda adalah spesies baru atau merupakan Au. afarensis.Australopithecus prometheus, atau dikenal sebagai Si Kaki Kecil baru-baru ini telah dihitung berasal dari 3,67 juta tahun yang lalu melalui teknik penanggalan baru, menjadikan genus Australopithecus berasal dari waktu yang sama dengan afarensis. Jempol kaki Si Kaki Kecil yang dapat ditekuk melintang dari jari-jari yang lain, menunjukkan bahwa spesimen itu tampaknya dulu adalah pemanjat yang baik. Mengingat keberadaan predator malam di wilayah tersebut, diperkirakan bahwa ia membangun tempat bersarang di malam hari di pepohonan dengan cara yang mirip dengan simpanse dan gorila.
Evolusi pada genus Homo
Representasi dari genus Homo yang paling awal terdokumentasi adalah Homo habilis, yang berevolusi sekitar 2,8 juta tahun yang lalu, dan dapat dikatakan sebagai spesies paling awal yang memiliki bukti positif tentang penggunaan alat-alat batu. Otak hominin awal tersebut memiliki ukuran yang hampir sama dengan simpanse, meskipun ada dugaan bahwa ini adalah masa di mana gen SRGAP2 manusia berlipat ganda, menghasilkan jaringan yang lebih cepat pada korteks frontal. Selama jutaan tahun berikutnya, proses ensefalisasi dengan pesat terjadi, dan dengan hadirnya Homo erectus dan Homo ergaster dalam rekaman fosil, kapasitas tengkorak telah berlipat ganda menjadi 850 cm3. (Peningkatan ukuran otak manusia seperti itu sama dengan setiap generasi memiliki 125.000 neuron lebih banyak dari orang tua mereka). Diyakini bahwa H. erectus dan H. ergaster adalah yang pertama kali menggunakan api dan peralatan yang kompleks, dan merupakan yang pertama dari garis hominin yang meninggalkan Afrika, menyebar ke seluruh Afrika, Asia, dan Eropa antara 1,3 hingga 1,8 juta tahun yang lalu.


|
−10 —
–
−9 —
–
−8 —
–
−7 —
–
−6 —
–
−5 —
–
−4 —
–
−3 —
–
−2 —
–
−1 —
–
0 —
|
|
|
||||||||||||||||||
Berdasarkan teori terkini mengenai asal-usul manusia modern di Afrika, manusia modern berevolusi di Afrika kemungkinan dari H. heidelbergensis, H. rhodesiensis, atau H. antecessor dan bermigrasi ke luar dari benua tersebut sekitar 50.000 hingga 100.000 tahun yang lalu, dan secara bertahap menggantikan populasi lokal H. erectus, hominin Denisova, H. floresiensis, H. luzonensis, dan H. neanderthalensis.Homo sapiens purba, cikal bakal manusia modern secara anatomis, berevolusi pada masa Paleolitik Tengah antara 400.000 hingga 250.000 tahun yang lalu. Bukti DNA terbaru menunjukkan bahwa beberapa haplotipe yang berasal dari Neanderthal ada di antara semua populasi non-Afrika, dan Neanderthal serta hominin lain, seperti Denisovan, mungkin telah menyumbangkan hingga 6% genom mereka kepada manusia masa kini, yang mengisyaratkan adanya perkawinan silang terbatas di antara spesies-spesies ini. Transisi menuju modernitas perilaku dengan perkembangan budaya simbolis, bahasa, dan teknologi litik khusus terjadi sekitar 50.000 tahun yang lalu, menurut beberapa antropolog, meskipun ada pula yang berpendapat bahwa perubahan perilaku secara bertahap terjadi dalam rentang waktu yang lebih lama.
Homo sapiens adalah satu-satunya spesies yang masih ada dari genusnya, Homo. Meskipun beberapa spesies Homo (yang telah punah) bisa jadi merupakan nenek moyang Homo sapiens, banyak, mungkin sebagian besar diperkirakan adalah "sepupu" dari Homo sapiens yang telah berspesiesisasi dari garis keturunan hominin. Belum ada konsensus mengenai kelompok mana yang harus dianggap sebagai spesies yang terpisah dan mana yang harus menjadi subspesies; hal ini mungkin disebabkan oleh kelangkaan fosil atau perbedaan kecil yang digunakan untuk mengklasifikasikan spesies dalam genus Homo. Teori pompa Sahara (yang mendeskripsikan kalau gurun Sahara terkadang-kadang “basah” sehingga bisa dilalui) memberikan satu penjelasan yang memungkinkan tentang variasi awal dalam genus Homo.
Berdasarkan bukti arkeologi dan paleontologi, telah memungkinkan untuk menyimpulkan, sampai batas tertentu, praktik pola makan kuno dari berbagai spesies Homo dan mempelajari peran pola makan dalam evolusi fisik dan perilaku dalam Homo.
Beberapa antropolog dan arkeolog menganut teori bencana Toba, yang menyatakan bahwa letusan dahsyat Danau Toba di Pulau Sumatera, Indonesia, sekitar 70.000 tahun yang lalu telah menimbulkan dampak global, menewaskan sebagian besar manusia dan menciptakan hambatan populasi yang mempengaruhi warisan genetik semua manusia saat ini. Namun, bukti genetik dan arkeologi untuk hal ini masih dipertanyakan.
H. habilis dan H. gautengensis
Homo habilis hidup dari sekitar 2,8 hingga 1,4 juta tahun lalu. Spesies ini berevolusi di Afrika Selatan dan Timur pada masa Pliosen Akhir atau Pleistosen Awal, 2,5-2 juta tahun yang lalu, ketika ia berpisah dari australopithecines dengan perkembangan gigi geraham yang lebih kecil dan otak yang lebih besar. Salah satu hominin pertama yang diketahui, membuat peralatan dari batu dan mungkin tulang binatang, yang menyebabkan namanya menjadi homo habilis (bahasa Latin yang berarti 'manusia praktis') yang diberikan oleh penemunya, Louis Leakey. Beberapa ilmuwan telah mengusulkan untuk memindahkan spesies ini dari Homo ke Australopithecus karena morfologi kerangkanya yang lebih beradaptasi untuk hidup di pepohonan daripada berjalan dengan dua kaki seperti hominin yang lebih baru.
Pada bulan Mei 2010, sebuah spesies baru, Homo gautengensis, ditemukan di Afrika Selatan.
H. rudolfensis dan H. georgicus
Ini adalah usulan nama spesies untuk fosil-fosil yang berasal dari sekitar 1,9-1,6 juta tahun yang lalu, yang hubungannya dengan Homo habilis masih belum jelas.
- Homo rudolfensis merujuk pada satu tengkorak yang tidak lengkap dari Kenya. Para ilmuwan berpendapat bahwa ini adalah spesimen Homo habilis, namun hal ini belum dikonfirmasi.
- Homo georgicus, dari Georgia, mungkin merupakan bentuk peralihan antara Homo habilis dan Homo erectus, atau subspesies Homo erectus.
H. ergaster dan H. erectus
Fosil pertama Homo erectus ditemukan oleh seorang dokter Belanda, Eugene Dubois, pada tahun 1891 di pulau Jawa, Indonesia. Dia awalnya menamai fosil tersebut Anthropopithecus erectus (1892-1893, pada saat itu dianggap sebagai fosil primata mirip simpanse) dan Pithecanthropus erectus (1893-1894, kemudian berubah pikiran karena morfologinya yang dianggap berada di tengah-tengah antara manusia dan kera). Bertahun-tahun kemudian, pada abad ke-20, dokter dan ahli paleoantropologi Jerman, Franz Weidenreich (1873-1948), membandingkan secara rinci karakter-karakter Manusia Jawa karya Dubois, yang saat itu diberi nama Pithecanthropus erectus, dengan karakter-karakter Manusia Peking, yang saat itu diberi nama Sinanthropus pekinensis. Weidenreich menyimpulkan pada tahun 1940 bahwa karena kemiripan anatomi mereka dengan manusia modern, maka perlu untuk mengumpulkan semua spesimen dari Jawa dan Cina dalam satu spesies dari genus Homo, yaitu spesies H. erectus.
Homo erectus hidup dari sekitar 1,8 juta tahun hingga sekitar 70.000 tahun yang lalu—yang mengindikasikan bahwa mereka mungkin dimusnahkan oleh bencana Toba; namun, H. floresiensis yang berada di dekatnya berhasil selamat. Fase awal H. erectus, dari 1,8 hingga 1,25 juta tahun lalu, dianggap oleh beberapa peneliti sebagai spesies yang terpisah, H. ergaster, atau sebagai H. erectus ergaster, sebuah subspesies dari H. erectus. Banyak ahli paleoantropologi sekarang menggunakan istilah Homo ergaster untuk bentuk-bentuk non-Asia dari kelompok ini, dan mencadangkan H. erectus hanya untuk fosil-fosil yang ditemukan di Asia dan memenuhi persyaratan kerangka dan gigi tertentu yang sedikit berbeda dari H. ergaster.
Di Afrika pada masa Pleistosen Awal, 1,5–1 juta tahun yang lalu, beberapa populasi Homo habilis diperkirakan telah berevolusi dengan otak yang lebih besar dan membuat peralatan batu yang lebih rumit; perbedaan-perbedaan ini dan perbedaan-perbedaan lainnya sudah cukup bagi para ahli antropologi untuk mengklasifikasikan mereka sebagai spesies baru, Homo erectus—di Afrika. Evolusi lutut yang mengunci dan pergerakan foramen magnum dianggap sebagai penyebab perubahan populasi yang lebih besar. Spesies ini juga mungkin telah menggunakan api untuk memasak daging. Richard Wrangham mencatat bahwa Homo tampaknya telah menjadi penghuni tanah, dengan berkurangnya panjang usus, gigi yang lebih kecil, dan "otak [membengkak] hingga ukurannya yang saat ini sangat tidak efisien dalam penggunaan bahan bakar", dan berhipotesis bahwa kendali atas api dan memasak, yang menghasilkan peningkatan nilai gizi, merupakan adaptasi utama yang memisahkan Homo dari Australopithecines yang tidur di pohon.
H. cepranensis and H. antecessor
Kedua spesies ini diusulkan sebagai spesies peralihan antara H. erectus dan H. heidelbergensis.
- H. antecessor diketahui dari fosil-fosil dari Spanyol dan Inggris yang berumur 1,2 juta-500 ribu tahun yang lalu.
- H. cepranensis mengacu pada satu tempurung tengkorak dari Italia, yang diperkirakan berusia sekitar 800.000 tahun.
H. heidelbergensis
H. heidelbergensis ("Manusia Heidelberg") hidup dari sekitar 800.000 hingga 300.000 tahun yang lalu. Juga diusulkan sebagai Homo sapiens heidelbergensis atau Homo sapiens paleohungaricus.
H. rhodesiensis, dan kranium Gawis
- H. rhodesiensis, diperkirakan hidup 300.000-125.000 tahun yang lalu. Sebagian besar peneliti saat ini menempatkan Manusia Rhodesia dalam kelompok Homo heidelbergensis, meskipun sebutan lain seperti Homo sapiens purba dan Homo sapiens rhodesiensis telah diusulkan.
- Pada bulan Februari 2006, sebuah fosil, tengkorak kepala Gawis, ditemukan yang mungkin merupakan spesies peralihan antara H. erectus dan H. sapiens atau salah satu dari sekian banyak evolusi yang berujung buntu. Tengkorak dari Gawis, Ethiopia, diyakini berasal dari masa 500.000-250.000 tahun yang lalu. Hanya rincian ringkasan yang diketahui, dan para penemunya belum merilis studi yang telah ditinjau oleh peneliti lain. Ciri-ciri wajah manusia Gawis mengisyaratkan bahwa ia adalah spesies peralihan atau contoh dari "manusia Bodo" perempuan.
Neanderthal dan Denisovan

Homo neanderthalensis, atau disebut juga sebagai Homo sapiens neanderthalensis, hidup di Eropa dan Asia dari 400.000 hingga sekitar 28.000 tahun yang lalu. Terdapat sejumlah perbedaan anatomi yang jelas antara manusia modern secara anatomis (AMH) dan spesimen Neanderthal, yang sebagian besar berkaitan dengan adaptasi Neanderthal yang lebih unggul terhadap lingkungan dingin. Rasio perbandingan antara alas dan volume Neanderthal bahkan lebih rendah dibandingkan dengan populasi Inuit modern, yang mengindikasikan retensi panas tubuh yang lebih baik.
Neanderthal juga memiliki otak yang jauh lebih besar, seperti yang ditunjukkan dari hasil endokopi otak, sehingga menimbulkan keraguan akan inferioritas intelektual mereka terhadap manusia modern. Namun, massa tubuh Neanderthal yang lebih tinggi mungkin membutuhkan massa otak yang lebih besar untuk kontrol tubuh. Selain itu, penelitian terbaru oleh Pearce, Stringer, dan Dunbar telah menunjukkan perbedaan penting dalam arsitektur otak. Ukuran yang lebih besar dari ruang orbital dan lobus oksipital Neanderthal menunjukkan bahwa mereka memiliki ketajaman penglihatan yang lebih baik daripada manusia modern, yang berguna pada saat kondisi cahaya yang lebih redup pada masa glasial di Eropa.
Manusia purba Neanderthal kemungkinan memiliki kapasitas otak yang lebih sedikit untuk fungsi sosial. Dengan menyimpulkan ukuran kelompok sosial dari volume endokranial (dikurangi ukuran lobus oksipital) menunjukkan bahwa pada setiap kelompok Neanderthal mungkin terbatas pada 120 individu, dibandingkan dengan 144 hubungan yang mungkin terjadi pada manusia modern. Kelompok sosial yang lebih besar dapat mengimplikasikan bahwa manusia modern memiliki risiko perkawinan sedarah yang lebih kecil di dalam kelompoknya, perdagangan di wilayah yang lebih luas (dikonfirmasi melalui distribusi alat-alat batu), dan penyebaran inovasi sosial dan teknologi yang lebih cepat. Semua hal ini bisa jadi berkontribusi pada Homo sapiens modern yang menggantikan populasi Neanderthal pada 28.000 tahun sebelum sekarang.
Bukti awal dari pengurutan DNA mitokondria menunjukkan bahwa tidak ada aliran gen yang signifikan antara H. neanderthalensis dan H. sapiens, dan bahwa keduanya adalah spesies terpisah yang memiliki nenek moyang yang sama sekitar 660.000 tahun yang lalu. Namun, pengurutan genom Neanderthal pada tahun 2010 mengindikasikan bahwa Neanderthal benar-benar melakukan perkawinan silang dengan manusia modern secara anatomis sekitar 45.000-80.000 tahun yang lalu, sekitar waktu manusia modern bermigrasi keluar dari Afrika, tetapi sebelum mereka menyebar ke seluruh Eropa, Asia, dan tempat lain. Pengurutan genetik kerangka manusia yang berasal dari 40.000 tahun yang lalu dari Rumania menunjukkan bahwa 11% dari genomnya adalah Neanderthal, menyiratkan bahwa individu tersebut memiliki nenek moyang Neanderthal 4-6 generasi sebelumnya, di samping kontribusi dari perkawinan silang lebih awal di Timur Tengah. Meskipun populasi Rumania hasil kawin silang ini tampaknya bukan nenek moyang manusia modern, temuan ini mengindikasikan bahwa kawin silang antara kedua spesies terjadi berulang kali.
Semua manusia modern non-Afrika memiliki sekitar 1% hingga 4% (atau 1,5% hingga 2,6% menurut data yang lebih baru) DNA yang berasal dari Neanderthal. Temuan ini konsisten dengan penelitian terbaru yang mengindikasikan bahwa perbedaan beberapa alel manusia berasal dari satu juta tahun yang lalu, meskipun interpretasi ini masih dipertanyakan. Neanderthal dan AMH Homo sapiens mungkin telah hidup berdampingan di Eropa selama 10.000 tahun, di mana populasi AMH meledak, jauh melebihi jumlah Neanderthal, sehingga mengalahkan mereka dalam hal jumlah.
Pada tahun 2008, para arkeolog yang bekerja di situs Gua Denisova di Pegunungan Altai, Siberia, menemukan sebuah fragmen tulang kecil dari jari kelima seorang anggota remaja dari spesies manusia lain, Denisovan. Berbagai artefak, termasuk gelang, ditemukan saat penggalian di gua tersebut pada tingkat yang sama dan memiliki usia karbon sekitar 40.000 tahun sebelum masehi. Dikarenakan iklim Gua Denisova yang dingin, DNA di dalam fragmen fosil itu bertahan dengan sangat baik, sehingga mtDNA maupun DNA nuklir-nya berhasil diurutkan.
Meskipun titik waktu divergensi mtDNA-nya secara tak terduga berada jauh, urutan genom lengkap menunjukkan bahwa Denisova berasal dari garis keturunan yang sama dengan Neanderthal, dan keduanya berbeda tak lama setelah garis keturunan mereka berpisah dari garis keturunan yang memunculkan manusia modern. Manusia modern diketahui telah hidup secara tumpang tindih dengan Neanderthal di Eropa dan Timur Dekat selama lebih dari 40.000 tahun, dan penemuan ini meningkatkan kemungkinan bahwa Neanderthal, Denisova, dan manusia modern mungkin telah hidup berdampingan dan kawin silang. Eksistensi cabang relasi yang jauh dari manusia ini menciptakan gambaran yang jauh lebih kompleks tentang umat manusia selama masa Pleistosen Akhir ketimbang yang diperkirakan sebelumnya. Telah ditemukan pula bukti bahwa sebanyak 6% DNA dari beberapa orang Melanesia modern berasal dari Denisovan, yang mengindikasikan adanya perkawinan silang yang terbatas di Asia Tenggara.
Sejumlah alel yang diperkirakan berasal dari Neanderthal dan Denisovan telah diidentifikasi pada beberapa lokus genetik dalam genom manusia modern di luar Afrika. Haplotipe HLA dari Denisovan dan Neanderthal mewakili lebih dari separuh alel HLA manusia modern yang natif di Eurasia, yang mengindikasikan adanya seleksi positif yang kuat untuk alel-alel yang terintroduksi ini. Corinne Simoneti di Vanderbilt University, di Nashville dan timnya telah menemukan dari catatan medis 28.000 orang keturunan Eropa bahwa keberadaan segmen DNA Neanderthal dapat dikaitkan dengan tingkat depresi yang lebih tinggi.
Aliran gen dari populasi Neanderthal ke manusia modern tidak hanya satu arah. Sergi Castellano dari Max Planck Institute for Evolutionary Anthropology melaporkan pada tahun 2016 bahwa meskipun genom Denisovan dan Neanderthal memiliki hubungan yang lebih dekat satu sama lain dibandingkan dengan kita, genom Neanderthal Siberia menunjukkan kemiripan yang lebih besar dengan gen manusia modern dibandingkan dengan populasi Neanderthal Eropa. Hal ini menunjukkan bahwa populasi Neanderthal kawin silang dengan manusia modern sekitar 100.000 tahun yang lalu, mungkin di suatu tempat di Timur Dekat.
Studi terhadap fosil seorang anak Neanderthal di Gibraltar menunjukkan bahwa dari perkembangan otak dan pertumbuhan gigi dapat disimpulkan bahwa anak-anak Neanderthal kemungkinan lebih cepat dewasa dibandingkan dengan Homo sapiens.
H. floresiensis

H. floresiensis, yang hidup sekitar 190.000 hingga 50.000 tahun sebelum sekarang, dijuluki hobbit karena ukurannya yang kecil, mungkin akibat dari kekerdilan insular.H. floresiensis cukup mencolok baik dari segi ukuran maupun usianya, dan merupakan contoh spesies terbaru dari genus Homo yang menunjukkan ciri-ciri turunan yang tidak dimiliki oleh manusia modern. Dengan kata lain, H. floresiensis memiliki nenek moyang yang sama dengan manusia modern, namun terpisah dari garis keturunan manusia modern dan mengikuti jalur evolusi yang berbeda. Temuan utama dari H. floresiensis adalah sebuah kerangka yang diyakini sebagai seorang perempuan berusia sekitar 30 tahun. Ditemukan pada tahun 2003, kerangka ini diperkirakan berasal dari masa sekitar 18.000 tahun yang lalu. Wanita yang masih hidup itu diperkirakan memiliki tinggi satu meter, dengan volume otak hanya 380 cm3 (tergolong kecil untuk simpanse dan kurang dari sepertiga dari rata-rata H. sapiens yang volume otaknya berukuran rata-rata 1.400 cm3).
Namun, sedang terdapat perdebatan tentang apakah H. floresiensis memang merupakan spesies yang terpisah atau tidak. Beberapa ilmuwan berpendapat bahwa H. floresiensis adalah H. sapiens modern dengan kekerdilan patologis. Hipotesis ini didukung sebagiannya karena beberapa manusia modern yang tinggal di Flores, pulau di Indonesia di mana kerangka itu ditemukan, bertubuh kerdil. Hal ini, ditambah dengan kekerdilan patologis, dapat menyebabkan manusia dengan tubuh yang sangat kecil. Argumen besar lainnya yang menentang H. floresiensis sebagai spesies yang terpisah adalah fakta bahwa spesies ini ditemukan dengan alat-alat yang hanya diasosiasikan dengan H. sapiens.
Akan tetapi, hipotesis kekerdilan patologis gagal menjelaskan fitur anatomi tambahan yang tidak seperti manusia modern (berpenyakit atau tidak) pada H. floresiensis, tetapi mirip dengan anggota purba dari genus Homo. Selain fitur tengkorak, fitur-fitur lain termasuk bentuk tulang di pergelangan tangan, lengan bawah, bahu, lutut, dan kaki. Selain itu, hipotesis ini gagal menjelaskan penemuan beberapa contoh individu dengan ciri-ciri yang sama, yang mengindikasikan bahwa ciri-ciri tersebut umum ditemukan pada populasi yang besar dari spesies tersebut, dan tidak terbatas pada satu individu.
Pada tahun 2016, fosil gigi dan sebagian rahang dari hominin yang dianggap sebagai nenek moyang H. floresiensis ditemukan di Mata Menge, sekitar 74 km (46 mil) dari Liang Bua. Fosil-fosil tersebut berasal dari sekitar 700.000 tahun yang lalu dan dicatat oleh arkeolog Australia, Gerrit van den Bergh, sebagai fosil yang lebih kecil daripada fosil-fosil yang ditemukan kemudian.
H. luzonensis
Sejumlah kecil spesimen dari pulau Luzon, yang berasal dari 50.000 hingga 67.000 tahun yang lalu, baru-baru ini oleh para penemunya, berdasarkan karakteristik gigi, ditetapkan sebagai spesies manusia baru, H. luzonensis.
H. sapiens
H. sapiens (kata sifat sapiens dalam bahasa Latin berarti "bijaksana" atau "cerdas") muncul di Afrika sekitar 300.000 tahun yang lalu, kemungkinan besar berasal dari H. heidelbergensis atau garis keturunan terkait. Pada September 2019, para ilmuwan melaporkan penentuan terkomputerisasi, berdasarkan 260 pemindaian CT, bentuk tengkorak virtual dari nenek moyang manusia terakhir yang sama dengan manusia modern/H. sapiens, yang mewakili manusia modern paling awal, dan menunjukkan bahwa manusia modern muncul antara 260.000 hingga 350.000 tahun yang lalu melalui penggabungan populasi di Afrika Timur dan Afrika Selatan.
Antara 400.000 tahun yang lalu hingga periode interglasial kedua pada Pleistosen Tengah, sekitar 250.000 tahun yang lalu, tren perluasan volume tengkorak dan elaborasi teknologi alat batu berkembang, sehingga memberi bukti adanya transisi dari H. erectus ke H. sapiens. Bukti langsung menunjukkan adanya migrasi H. erectus ke luar Afrika, kemudian spesiasi lebih lanjut dari H. sapiens dari H. erectus di Afrika. Migrasi berikutnya dari H. sapiens (baik di dalam maupun di luar Afrika) pada akhirnya menggantikan H. erectus yang telah terdispersi sebelumnya. Teori migrasi dan asal-usul ini biasanya disebut sebagai "hipotesis asal tunggal baru-baru ini" atau teori "keluar dari Afrika". H. sapiens melakukan kawin silang dengan manusia purba baik di Afrika maupun di Eurasia, di Eurasia terutama dengan Neanderthal dan Denisovan.
Teori bencana Toba, yang mendalilkan adanya hambatan populasi H. sapiens sekitar 70.000 tahun yang lalu, menjadi kontroversi sejak pertama kali diajukan pada tahun 1990-an dan pada tahun 2010-an hanya mendapat sedikit dukungan. Variabilitas genetik manusia yang khas telah muncul sebagai hasil dari efek pendiri, oleh percampuran purba dan oleh tekanan evolusi baru-baru ini.
Perubahan-perubahan anatomi
Evolusi manusia sejak berpisahnya dari nenek moyang bersama manusia dan simpanse yang terakhir, dicirikan oleh sejumlah perubahan morfologi, perkembangan, fisiologi, perilaku, dan lingkungan. Evolusi lingkungan (kultural) yang ditemukan jauh setelahnya selama era Pleistosen memainkan peran penting dalam evolusi manusia yang diobservasi melalui transisi manusia di antara sistem-sistem subsistensi. Yang paling signifikan dari adaptasi ini adalah bipedalisme, peningkatan ukuran otak, ontogeni yang memanjang (kehamilan dan masa bayi), dan penurunan dimorfisme seksual. Perubahan morfologi signifikan lainnya termasuk evolusi kekuatan dan presisi cengkeraman, yang pertama kali terjadi pada H. erectus.
Bipedalisme
Bipedalisme atau berjalan dengan dua kaki adalah adaptasi dasar hominid dan dianggap sebagai penyebab utama di balik serangkaian perubahan kerangka yang terjadi oleh semua hominid bipedal. Hominin paling awal, yang diduga primitif bipedalisme, diyakini sebagai Sahelanthropus atau Orrorin, yang keduanya muncul sekitar 6 sampai 7 juta tahun yang lalu. Sedangkan gorila dan simpanse, yang mengikut sertakan tangan mereka untuk berjelan, berpisah dari garis hominin sepanjang periode yang meliputi waktu yang sama, sehingga Sahelanthropus atau Orrorin bisa jadi merupakan nenek moyang bersama terakhir kita. Ardipithecus, yang berjalan secara penuh dengan kaki dua, muncul sekitar 5,6 juta tahun lalu.

Bipedal awal akhirnya berevolusi menjadi australopithecines dan kemudian menjadi genus Homo. Ada beberapa teori tentang nilai adaptasi bipedalisme. Ada kemungkinan bahwa bipedalisme disukai karena membebaskan tangan untuk menjangkau dan membawa makanan, menghemat energi selama pergerakan, memungkinkan lari jarak jauh dan berburu, memberikan bidang penglihatan yang lebih baik, dan membantu menghindari hipertermia dengan mengurangi luas permukaan yang terpapar sinar matahari langsung; fitur-fitur yang semuanya menguntungkan untuk berkembang di lingkungan sabana dan hutan baru yang tercipta sebagai hasil pengangkatan Lembah Celah Afrika Timur yang berbanding terbalik dengan habitat hutan tertutup sebelumnya.. Sebuah studi tahun 2007 mendukung hipotesis bahwa berjalan dengan dua kaki, atau bipedalisme, berkembang karena membutuhkan lebih sedikit energi daripada berjalan dengan kaki dan lengan. Namun, penelitian terbaru menunjukkan bahwa bipedalisme tanpa kemampuan menggunakan api tidak akan memungkinkan penyebaran global. Perubahan dalam gaya berjalan ini menyebabkan pemanjangan kaki secara proporsional jika dibandingkan dengan panjang lengan, yang memendek akibat hilangnya kebutuhan untuk bergelayutan dari cabang pohon satu ke cabang pohon lainnya. Perubahan lainnya adalah bentuk jempol kaki. Studi terbaru menunjukkan bahwa australopithecine masih menjalani sebagian hidupnya di pepohonan sebagai hasil dari mempertahankan jempol kaki yang dapat menggenggam. Hal ini secara progresif hilang pada habilin.
Secara anatomis, evolusi bipedalisme diiringi dengan sejumlah besar perubahan kerangka, tidak terbatas hanya pada bagian tungkai dan panggul, tetapi juga pada tulang belakang, telapak kaki dan pergelangan kaki, serta tengkorak. Tulang paha berevolusi ke posisi yang sedikit lebih angular untuk memindahkan pusat gravitasi ke arah pusat geometris tubuh. Sendi lutut dan pergelangan kaki menjadi semakin kokoh untuk menopang peningkatan berat badan dengan lebih baik. Untuk mendukung peningkatan berat pada tulang punggung dalam posisi tegak, kolom vertebra manusia berubah menjadi berbentuk S dan vertebra lumbar berubah menjadi lebih pendek dan lebih lebar. Pada kaki, jempol kaki berpindah ke posisi sejajar dengan jari-jari kaki yang lain untuk membantu pergerakan ke depan. Lengan dan lengan bawah memendek relatif terhadap kaki sehingga lebih mudah untuk berlari. Foramen magnum berpindah ke bawah tengkorak dan lebih ke anterior.
Perubahan paling signifikan terjadi di daerah panggul, di mana bilah iliaka yang panjang menghadap ke bawah memendek dan melebar sebagai keperluan untuk menjaga kestabilan pusat gravitasi saat berjalan; hominid yang bipedal memiliki panggul yang lebih pendek tetapi lebih luas seperti mangkuk karena hal ini. Kelemahannya, saluran lahir kera bipedal lebih kecil daripada kera yang berjalan menggunakan kaki dan tangan, meskipun telah terjadi pelebaran dibandingkan dengan australopithecine dan manusia modern, sehingga memungkinkan lewatnya bayi yang baru lahir karena peningkatan ukuran kranial. Hal ini terbatas pada bagian atas, karena peningkatan lebih lanjut dapat menghambat gerakan bipedal yang normal.
Evolusi yang terjadi pada panggul yang menjadi lebih pendek dan saluran lahir yang menjadi lebih kecil terjadi sebagai syarat bipedalisme yang berefek signifikan pada proses kelahiran manusia modern, yang mana jauh lebih sulit dibandingkan primata lainnya. Selama kelahiran manusia, karena variasi ukuran daerah panggul, kepala janin harus berada dalam posisi melintang (dibandingkan dengan ibu) selama melintasi saluran kelahiran dan berputar sekitar 90 derajat saat keluar. Saluran kelahiran yang lebih kecil menjadi faktor pembatas untuk peningkatan ukuran otak pada manusia purba dan mendorong periode kehamilan yang lebih pendek yang mengarah pada ketidakdewasaan relatif pada keturunan manusia, yang tidak dapat berjalan jauh sebelum 12 bulan dan memiliki perkembangan fisiologi atau somatis yang lebih lambat dibandingkan dengan primata lain, yang dapat berjalan pada usia yang jauh lebih dini. Pertumbuhan otak yang meningkat setelah lahir dan meningkatnya ketergantungan anak-anak pada ibu mereka memiliki efek besar pada siklus reproduksi wanita, yang mengakibatkan lebih banyaknya kemunculan pengasuhan anak yang dibantu oleh kerabatnya atau orang lain pada manusia jika dibandingkan dengan hominid lainnya. Kematangan seksual manusia yang tertunda juga menyebabkan evolusi menopause, dengan salah satu penjelasannya adalah hipotesis nenek, yakni wanita lanjut usia akan dapat meneruskan gen mereka dengan lebih baik dengan merawat keturunan dari anak perempuan mereka, dibandingkan dengan memiliki lebih banyak anak sendiri.
Ensefalisasi


Spesies manusia lambat laun mengembangkan otak yang jauh lebih besar daripada primata lainnya-biasanya, 1.330 cm3 (81 cu in) pada manusia modern, hampir tiga kali ukuran otak simpanse atau gorila. Setelah periode stasis dengan Australopithecus anamensis dan Ardipithecus, spesies yang memiliki otak lebih kecil sebagai akibat dari lokomosi bipedal mereka, pola ensefalisasi bermula pada Homo habilis, yang otaknya 600 cm3 (37 cu in) sedikit lebih besar daripada simpanse. Evolusi ini berlanjut pada Homo erectus dengan 800–1,100 cm3 (48,8190–0,0671 cu in), dan mencapai maksimum pada Neanderthal dengan 1.200–1,900 cm3 (73,2285–0,1159 cu in), bahkan lebih besar dari Homo sapiens modern. Peningkatan otak ini terwujud selama pertumbuhan otak pascakelahiran, jauh melebihi kera lainnya (heterokroni). Hal ini juga memungkinkan periode pembelajaran sosial dan akuisisi bahasa yang lebih lama pada manusia remaja, yang dimulai sejak 2 juta tahun yang lalu. Ensefalisasi mungkin disebabkan oleh ketergantungan pada makanan padat kalori yang sulit diperoleh.
Lebih lanjut, perubahan dalam struktur otak manusia kemungkinan bahkan lebih signifikan daripada peningkatan ukurannya.

Lobus temporal, yang berisi pusat-pusat pemrosesan bahasa, telah meningkat secara tidak proporsional, seperti halnya korteks prefrontal, yang telah dikaitkan dengan pengambilan keputusan yang kompleks dan memoderasi perilaku sosial. Ensefalisasi telah dikaitkan dengan meningkatnya pati dan daging dalam makanan, namun sebuah studi meta tahun 2022 meragukan peran dari daging. Faktor lainnya adalah perkembangan memasak, dan telah diajukan bahwa kecerdasan meningkat sebagai respons terhadap peningkatan kebutuhan untuk memecahkan masalah sosial karena masyarakat manusia menjadi lebih kompleks. Perubahan morfologi tengkorak, seperti mandibula yang mengecil dan pelekatan otot mandibula, memungkinkan lebih banyak ruang bagi otak untuk berkembang.
Peningkatan volume neokorteks juga mencakup peningkatan pesat dalam ukuran serebelum. Fungsinya secara tradisional dikaitkan dengan keseimbangan dan kontrol motorik halus, tetapi baru-baru ini ditemukan juga berkaitan dengan kemampuan bicara dan kognisi. Kera besar, termasuk hominid, memiliki serebelum yang lebih menonjol relatif terhadap neokorteks daripada primata lainnya. Diduga bahwa karena fungsinya sebagai kontrol sensorik-motorik dan pembelajaran aktivitas otot yang kompleks, serebelum mungkin telah mendasari adaptasi teknologi manusia, termasuk prakondisi bicara.
Manfaat langsung dari ensefalisasi untuk bertahan hidup sulit untuk dipahami, karena perubahan otak utama dari Homo erectus ke Homo heidelbergensis tidak disertai dengan perubahan besar dalam bidang teknologi. Diduga bahwa perubahan tersebut sebagian besar bersifat sosial dan perilaku, termasuk peningkatan kemampuan empati, peningkatan ukuran kelompok sosial, dan peningkatan fleksibilitas dalam berperilaku. Manusia memiliki kemampuan unik untuk memperoleh informasi melalui transmisi sosial dan beradaptasi dengan informasi tersebut. Bidang evolusi kultural yang sedang berkembang mengkaji perubahan sosiokultural manusia dari perspektif evolusi.

Dimorfisme seksual
Berkurangnya tingkat dimorfisme seksual pada manusia terlihat terutama pada penyusutan gigi taring pada laki-laki dibandingkan dengan spesies kera lainnya (kecuali owa) dan berkurangnya tonjolan alis dan fisik keseluruhan pada pria. Perubahan fisiologis utama lainnya yang berkaitan dengan seksualitas pada manusia adalah evolusi estrus terpendam. Manusia adalah satu-satunya hominoid yang betinanya subur sepanjang tahun dan tidak ada sinyal khusus kesuburan yang dihasilkan oleh tubuh (seperti pembengkakan genital atau perubahan mencolok dalam perilakunya kepada laki-laki selama estrus).
Meski demikian, sejumlah dimorfisme seksual tetap bertahan pada manusia, seperti dalam pendistribusian rambut tubuh dan lemak subkutan, dan dalam ukuran keseluruhan, laki-laki sekitar 15% lebih besar daripada perempuan. Perubahan-perubahan ini secara keseluruhan telah diartikan sebagai hasil dari meningkatnya penekanan pada pembentukan ikatan pasangan sebagai solusi yang memungkinkan untuk memenuhi meningkatnya keterlibatan orang tua pada anak-anak mereka karena masa pertumbuhannya yang menjadi lebih lama.
Oposisi ulnar

Oposisi ulnar yakni kontak antara ibu jari dan ujung jari kelingking dari tangan yang sama, adalah unik untuk genus Homo, termasuk Neanderthal, para hominin Sima de los Huesos, dan manusia modern secara anatomis. Pada primata lain, jari jempolnya berukuran pendek dan tidak dapat menyentuh jari kelingkingnya. Oposisi ulnar memfasilitasi cengkeraman presisi dan cengkeraman kekuatan pada tangan manusia, yang mendasari segala manipulasi yang terampil.
Perubahan-perubahan lain
Sejumlah perubahan lain juga mengkarakterisasi evolusi manusia, di antaranya peningkatan ketergantungan pada penglihatan ketimbang penciuman (bulbus olfaktorius yang amat tereduksi); periode perkembangan remaja yang lebih panjang dan ketergantungan bayi yang lebih tinggi; usus yang lebih kecil dan gigi yang kecil dan tidak selaras; metabolisme basal yang lebih cepat; hilangnya rambut tubuh; evolusi kelenjar keringat; perubahan bentuk arkade gigi dari bentuk u menjadi parabola; perkembangan dagu (hanya ditemukan di Homo sapiens); projeksi styloid; dan laring yang turun.
Sejarah pembelajaran
Sebelum Darwin
Kata homo, nama genus biologis dimana manusia termasuk di dalamnya, adalah bahasa Latin untuk "manusia". Kata ini awalnya dipilih oleh Carl Linnaeus dalam sistem klasifikasinya. Kata bahasa Inggris dari manusia, yaitu "human" berasal dari bahasa Latin humanus, bentuk kata sifat dari kata homo. Bahasa Latin "homo" berakar dari bahasa proto-Indo-Eropa dari kata *dhghem atau "bumi". Linnaeus dan ilmuwan lain pada masanya juga menganggap kera besar sebagai kerabat terdekat manusia berdasarkan kesamaan morfologi dan anatomi.
Darwin
Adanya kemungkinan keterkaitan antara manusia dengan kera-kera terdahulu melalui keturunan, baru menjadi jelas setelah tahun 1859 dengan diterbitkannya buku Charles Darwin, On the Origin of Species (Asal-usul Spesies), di mana ia berargumen tentang konsep evolusi spesies baru dari spesies yang lebih awal. Buku Darwin tersebut tidak membahas masalah evolusi manusia, hanya mengatakan bahwa "Cahaya akan dilemparkan pada asal-usul manusia dan sejarahnya."
Perdebatan pertama tentang sifat evolusi manusia muncul antara Thomas Henry Huxley dan Richard Owen. Huxley berargumen tentang evolusi manusia dari kera dengan mengilustrasikan banyak kesamaan dan perbedaan antara manusia dan kera lainnya, dan melakukannya secara khusus dalam bukunya di tahun 1863 yang berjudul Evidence as to Man's Place in Nature (Bukti tentang Tempat Manusia di Alam). Banyak pendukung awal Darwin (seperti Alfred Russel Wallace dan Charles Lyell) pada awalnya tidak setuju bahwa asal mula kapasitas mental dan kepekaan moral manusia dapat dijelaskan oleh seleksi alam, meskipun akhirnya keyakinan ini berubah. Darwin menerapkan teori evolusi dan seleksi seksual pada manusia dalam bukunya yang diterbitkan tahun 1871, The Descent of Man, and Selection in Relation to Sex (Kemunculan Manusia, dan Seleksi dalam Kaitannya dengan Seks).
Fosil-fosil pertama
Masalah utama pada abad ke-19 adalah kurangnya fosil perantara. Sisa-sisa Neanderthal ditemukan di sebuah tambang batu kapur pada tahun 1856, tiga tahun sebelum publikasi On the Origin of Species, dan fosil Neanderthal telah ditemukan di Gibraltar bahkan lebih awal, tetapi pada awalnya fosil ini diklaim sebagai sisa-sisa manusia modern yang telah mengalami beberapa jenis penyakit. Meskipun pada tahun 1891 ditemukan oleh Eugène Dubois dari apa yang sekarang disebut Homo erectus di Trinil, Jawa, baru pada tahun 1920-an lah fosil-fosil seperti itu baru ditemukan di Afrika, dan bukti-bukti tentang spesies perantara pun mulai terakumulasi. Pada tahun 1925, Raymond Dart mendeskripsikan Australopithecus africanus. Spesimen jenisnya adalah Bocah Taung, bayi australopithecine yang ditemukan di sebuah gua. Sisa-sisa anak tersebut adalah berupa tengkorak kecil yang terawetkan dengan sangat baik dan sebuah endocast otak.
Meskipun ukuran otaknya kecil (410 cm3), namun bentuknya bulat, tidak seperti simpanse dan gorila, dan lebih mirip otak manusia modern. Selain itu, spesimen tersebut memiliki gigi taring pendek, dan posisi foramen magnum (lubang di tengkorak tempat masuknya tulang belakang) adalah bukti dari kemampuan berjalan dengan dua kaki. Semua ciri-ciri ini meyakinkan Dart bahwa Bocah Taung adalah nenek moyang manusia bipedal, bentuk transisi antara kera dan manusia.
Fosil-fosil Afrika Timur

Selama tahun 1960-an dan 1970-an, ratusan fosil ditemukan di Afrika Timur di wilayah Ngarai Olduvai dan Danau Turkana. Pencarian ini dilakukan oleh keluarga Leakey, yaitu Louis Leakey dan istrinya Mary Leakey, dan kemudian anak mereka Richard dan menantu perempuannya Meave, dengan beberapa pemburu fosil dan paleoantropolog. Dari lapisan fosil Olduvai dan Danau Turkana, mereka menemukan sejumlah spesimen hominin awal, seperti spesies australopithecines dan spesies Homo, bahkan Homo erectus.
Penemuan-penemuan ini mengukuhkan Afrika sebagai tempat kelahiran umat manusia. Pada akhir 1970-an dan 1980-an, Ethiopia muncul sebagai titik panas baru paleoantropologi setelah penemuan "Lucy", anggota fosil paling lengkap dari spesies Australopithecus afarensis, yang ditemukan pada tahun 1974 oleh Donald Johanson di dekat Hadar di wilayah gurun Segitiga Afar di Ethiopia utara. Meskipun spesimen tersebut memiliki otak yang kecil, namun panggul dan tulang kakinya hampir identik fungsinya dengan manusia modern, yang menunjukkan dengan pasti bahwa hominin ini telah berjalan tegak. Lucy diklasifikasikan sebagai spesies baru, Australopithecus afarensis, yang dianggap lebih dekat hubungannya dengan genus Homo sebagai nenek moyang langsung, atau sebagai kerabat dekat dari nenek moyang yang tidak diketahui, dibandingkan hominid atau hominin lain yang diketahui dari rentang waktu awal ini; lihat istilah "hominid" dan "hominin". (Nama "Lucy" pada spesimen ini diambil dari lagu the Beatles "Lucy in the Sky with Diamonds", yang diputar dengan keras dan berulang-ulang di kamp selama penggalian). Kawasan Segitiga Afar kemudian mendatangkan lebih banyak penemuan fosil hominin, terutama yang ditemukan atau dideskripsikan oleh tim yang dipimpin oleh Tim D. White pada 1990-an, termasuk Ardipithecus ramidus dan Ardipithecus kadabba.
Pada tahun 2013, kerangka fosil Homo naledi, spesies hominin yang telah punah yang ditempatkan (sementara) ke dalam genus Homo, ditemukan di sistem Gua Rising Star, sebuah situs di wilayah Tempat Kelahiran Umat Manusia di Afrika Selatan di provinsi Gauteng dekat Johannesburg. Hingga September 2015, fosil dari setidaknya lima belas individu, berjumlah 1.550 spesimen, telah digali dari gua tersebut. Spesies ini dicirikan oleh massa tubuh dan perawakan yang mirip dengan populasi manusia bertubuh kecil, volume endokranial yang lebih kecil mirip dengan Australopithecus, dan morfologi kranial (bentuk tengkorak) yang mirip dengan spesies Homo awal. Anatomi kerangka memadukan fitur primitif yang dikenal dari australopithecines dengan fitur yang dikenal dari hominin awal. Individu-individu tersebut menunjukkan adanya tanda-tanda pembuangan yang disengaja ke dalam gua menjelang waktu kematian mereka. Fosil-fosil tersebut berasal dari 250.000 tahun yang lalu, yang berarti bukan merupakan nenek moyang langsung manusia, melainkan hidup pada waktu yang sama dengan kemunculan pertama manusia modern secara anatomis yang berotak lebih besar.
Revolusi Genetik
Revolusi genetik dalam studi evolusi manusia berawal ketika Vincent Sarich dan Allan Wilson mengukur kekuatan reaksi silang imunologis dari serum darah albumin di antara beberapa pasangan makhluk hidup, termasuk manusia dan kera Afrika (simpanse dan gorila). Kekuatan reaksinya dapat dinyatakan secara numerik sebagai jarak imunologis, yang ternyata berbanding lurus dengan jumlah perbedaan asam amino antara protein homolog pada spesies yang berbeda. Dengan membangun kurva kalibrasi dari ID pasangan-pasangan spesies dengan waktu pemisahan yang diketahui dalam catatan fosil, data tersebut dapat digunakan sebagai jam molekuler untuk mengestimasi waktu pemisahan dari pasangan-pasangan dengan catatan fosil yang lebih buruk atau tidak diketahui.
Dalam makalah seminal mereka tahun 1967 di Science, Sarich dan Wilson memperkirakan waktu pemisahan manusia dan kera adalah empat hingga lima juta tahun yang lalu, di mana saat itu interpretasi standar dari catatan fosil memberikan waktu pemisahan ini setidaknya 10 hingga 30 juta tahun yang lalu. Penemuan fosil berikutnya, terutama "Lucy", dan interpretasi ulang dari materi fosil yang lebih tua, terutama Ramapithecus, menunjukkan bahwa perkiraan yang lebih muda adalah yang tepat dan memvalidasi metode albumin.
Progres dalam pengurutan DNA, khususnya DNA mitokondria (mtDNA) dan kemudian DNA kromosom Y (Y-DNA) meningkatkan pemahaman tentang asal-usul manusia. Penerapan prinsip jam molekuler merevolusi studi evolusi molekuler.
Berdasarkan waktu pemisahan manusia dengan orangutan yang terjadi antara 10 dan 20 juta tahun lalu, studi awal jam molekuler menunjukkan bahwa ada sekitar 76 mutasi per generasi yang tidak diwariskan oleh anak-anak manusia dari orang tua mereka; bukti ini mendukung waktu pemisahan antara hominin dan simpanse seperti yang disebutkan di atas. Namun, penelitian pada 2012 di Islandia terhadap 78 anak dan orang tua mereka menunjukkan tingkat mutasi hanya 36 mutasi per generasi; data ini memperpanjang waktu pemisahan antara manusia dan simpanse ke periode yang lebih awal, lebih dari 7 juta tahun lalu (Ma). Penelitian tambahan dengan 226 keturunan populasi simpanse liar di delapan lokasi menunjukkan bahwa simpanse bereproduksi rata-rata pada usia 26,5 tahun; yang menunjukkan pemisahan manusia dengan simpanse terjadi antara 7 dan 13 juta tahun lalu. Dan data ini menunjukkan bahwa Ardipithecus (4,5 Ma), Orrorin (6 Ma), dan Sahelanthropus (7 Ma) semuanya mungkin berada pada garis keturunan hominid, dan bahkan pemisahan itu mungkin terjadi di luar wilayah Rengkahan Afrika Timur.
Lebih lanjut, analisis terhadap gen kedua spesies tersebut pada tahun 2006 membuktikan bahwa setelah nenek moyang manusia mulai terpisah dari simpanse, perkawinan antarspesies antara "proto-manusia" dan "proto-simpanse" tetap terjadi secara cukup rutin yang mengakibatkan perubahan gen-gen tertentu dalam lungkang gen baru:
- Perbandingan baru genom manusia dan simpanse menunjukkan bahwa setelah garis keturunan keduanya berpisah, mereka kemungkinan telah mulai melakukan kawin silang.... Temuan utamanya adalah bahwa kromosom X manusia dan simpanse tampaknya mengalami pemisahan sekitar 1,2 juta tahun lebih kini daripada kromosom lainnya.
Penelitian ini menunjukkan:
- Faktanya adalah terdapat dua pemisahan antara garis keturunan manusia dan simpanse, dengan yang pertama diikuti oleh perkawinan silang antara dua populasi spesies tersebut dan kemudian pemisahan kedua. Dugaan adanya hibridisasi telah mengejutkan para ahli paleoantropologi, yang tetap memperlakukan data genetik baru ini dengan amat serius.
Pencarian hominin paling awal
Pada tahun 1990-an, beberapa tim paleoantropolog bekerja di seantero Afrika untuk mencari bukti pemisahan paling awal garis keturunan hominin dari kera besar. Pada 1994, Meave Leakey menemukan Australopithecus anamensis. Namun, penemuan ini kemudian tenggelam oleh penemuan Ardipithecus ramidus oleh Tim D. White pada tahun 1995, yang memundurkan catatan fosil hingga 4,2 juta tahun silam.
Pada tahun 2000, Martin Pickford dan Brigitte Senut menemukan, di Perbukitan Tugen, Kenya, hominin bipedal berusia 6 juta tahun yang mereka beri nama Orrorin tugenensis. Dan pada tahun 2001, sebuah tim yang dipimpin oleh Michel Brunet menemukan tengkorak Sahelanthropus tchadensis yang berumur 7,2 hingga 20 juta tahun silam, dan Brunet berpendapat bahwa tengkorak tersebut adalah bipedal, dan oleh karena itu adalah hominin, yang merupakan hominid. (lihat istilah "hominid" dan hominin).
Penyebaran manusia


Para antropolog pada tahun 1980-an terpecah mengenai beberapa detail tentang halangan reproduksi dan penyebaran migrasi dari genus Homo. Belakangan, ilmu genetika digunakan untuk menyelidiki dan menyelesaikan masalah-masalah ini. Menurut teori pompa Sahara, bukti-bukti menunjukkan bahwa genus Homo bermigrasi keluar dari Afrika setidaknya tiga dan mungkin empat kali (misalnya Homo erectus, Homo heidelbergensis dan dua atau tiga kali untuk Homo sapiens). Bukti-bukti terbaru menunjukkan bahwa penyebaran ini terkait erat dengan periode perubahan iklim yang berfluktuasi.
Bukti terbaru menunjukkan bahwa manusia kemungkinan meninggalkan Afrika setengah juta tahun lebih awal dari yang diperkirakan sebelumnya. Sebuah tim gabungan Perancis-India telah menemukan artefak manusia di Bukit Siwalk di utara New Delhi yang berasal dari setidaknya 2,6 juta tahun yang lalu. Ini lebih awal dari temuan genus Homo paling awal sebelumnya di Dmanisi, di Georgia, yang berasal dari 1,85 juta tahun yang lalu. Meskipun kontroversial, perkakas yang ditemukan di sebuah gua di Tiongkok memperkuat argumen bahwa manusia menggunakan perkakas sejak 2,48 juta tahun yang lalu. Hal ini menunjukkan bahwa kelompok dengan tradisi perkakas "Pemotong" di Asia, yang ditemukan di Jawa dan Cina bagian utara mungkin telah pergi meninggalkan Afrika sebelum kemunculan kapak tangan Acheulian.
Penyebaran Homo sapiens modern
Hingga bukti genetik tersedia, ada dua model dominan untuk penyebaran manusia modern. Hipotesis multiregional mengusulkan bahwa genus Homo hanya terdiri dari satu populasi yang saling berkaitan seperti saat ini (bukan spesies yang terpisah), dan evolusinya terjadi di seluruh dunia secara terus menerus selama beberapa juta tahun terakhir. Model ini diusulkan pada tahun 1988 oleh Milford H. Wolpoff. Sebaliknya, model "keluar dari Afrika" mengusulkan bahwa H. sapiens modern berspesialisasi di Afrika baru-baru ini (yaitu, sekitar 200.000 tahun yang lalu) dan migrasi berikutnya melalui Eurasia mengakibatkan penggantian spesies Homo lainnya yang hampir lengkap. Model ini dikembangkan oleh Chris B. Stringer dan Peter Andrews.

Pengurutan mtDNA dan Y-DNA yang diambil sampelnya dari berbagai populasi pribumi menunjukkan informasi leluhur yang berkaitan dengan warisan genetik pria dan wanita, dan memperkuat teori "keluar dari Afrika" dan melemahkan pandangan evolusionisme multiregional. Perbedaan pohon genetik yang selaras diartikan sebagai pendukung model asal usul tunggal yang baru saja muncul.
Model "Keluar dari Afrika" dengan demikian telah memperoleh dukungan yang besar dari penelitian yang menggunakan DNA mitokondria wanita dan kromosom Y pria. Setelah menganalisis pohon silsilah yang dibangun dengan menggunakan 133 jenis mtDNA, para peneliti menyimpulkan bahwa semuanya adalah keturunan dari nenek moyang Afrika perempuan, yang dijuluki Hawa Mitokondria. Model "Keluar dari Afrika" juga didukung oleh fakta bahwa keragaman genetik mitokondria paling tinggi terdapat di antara populasi Afrika.
Sebuah studi yang ekstensif tentang keragaman genetik Afrika, yang dipimpin oleh Sarah Tishkoff, menemukan bahwa orang San memiliki keragaman genetik terbesar di antara 113 populasi berbeda yang dijadikan sampel, menjadikan mereka salah satu dari 14 "kelompok populasi leluhur". Penelitian ini juga menemukan kemungkinan asal muasal migrasi manusia modern di Afrika barat daya, dekat perbatasan pantai Namibia dan Angola. Namun, bagi arkeolog Richard Leakey, bukti fosil itu tidak cukup untuk menyelesaikan perdebatan tentang di mana tepatnya di Afrika, manusia modern pertama kali muncul. Studi haplogroups dalam DNA kromosom Y dan DNA mitokondria telah sebagian besar memperkuat asal usul Afrika yang baru-baru ini. Model ini juga sangat didukung oleh seluruh bukti dari DNA autosomal. Namun demikian, bukti adanya percampuran purba pada manusia modern, baik di Afrika, dan kemudian di seluruh Eurasia, belakangan telah dinyatakan oleh sejumlah penelitian.
Pengurutan terbaru dari genom Neanderthal dan Denisovan menunjukkan bahwa beberapa percampuran antara manusia modern dengan populasi ini telah terjadi. Semua kelompok manusia modern di luar Afrika memiliki 1-4% atau (menurut penelitian yang lebih baru) sekitar 1,5-2,6% alel Neanderthal dalam genom mereka, dan beberapa orang Melanesia memiliki tambahan 4-6% alel Denisovan. Hasil-hasil baru ini tidak bertentangan dengan model "keluar dari Afrika", kecuali dalam interpretasinya yang paling ketat, meskipun hasil-hasil ini membuat situasi menjadi lebih kompleks. Setelah pulih dari hambatan genetik yang menurut beberapa peneliti mungkin terkait dengan bencana gunung berapi supervolkano Toba, kelompok yang cukup kecil meninggalkan Afrika dan kawin silang dengan Neanderthal, mungkin di Timur Tengah, di padang rumput Eurasia atau bahkan di Afrika Utara sebelum kepergian mereka. Keturunan mereka yang masih didominasi Afrika menyebar untuk mengisi dunia. Sebagian kecil pada gilirannya kawin silang dengan Denisovan, mungkin di Asia Tenggara, sebelum mendiami Melanesia. Haplotipe HLA asal Neanderthal dan Denisovan telah diidentifikasi terdapat pada populasi Eurasia dan Oseania modern. Gen EPAS1 Denisovan juga telah ditemukan pada populasi Tibet. Studi genom manusia menggunakan pembelajaran mesin telah mengidentifikasi kontribusi genetik tambahan pada orang Eurasia dari populasi leluhur yang "tidak diketahui" yang berpotensi berhubungan dengan garis keturunan Neanderthal-Denisovan.
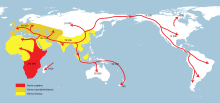
Masih terdapat perbedaan teori tentang apakah ada eksodus tunggal dari Afrika atau beberapa. Model penyebaran ganda melibatkan teori Penyebaran Selatan, yang telah mendapatkan dukungan dalam beberapa tahun terakhir dari bukti genetik, linguistik dan arkeologi. Dalam teori ini, ada penyebaran pesisir manusia modern dari wilayah Tanduk Afrika menyeberangi Bab el Mandib ke Yaman pada permukaan laut yang lebih rendah sekitar 70.000 tahun yang lalu. Kelompok ini membantu mengisi Asia Tenggara dan Oseania, menjelaskan penemuan situs-situs manusia purba di daerah-daerah ini jauh lebih awal daripada yang ada di Syam. Kelompok ini tampaknya bergantung pada sumber daya laut untuk kelangsungan hidup mereka.
Stephen Oppenheimer mengajukan gelombang kedua manusia mungkin kemudian menyebar melalui oasis Teluk Persia, dan pegunungan Zagros ke Timur Tengah. Secara alternatif, gelombang kedua ini mungkin datang melintasi Semenanjung Sinai ke Asia, dari tak lama setelah 50.000 tahun yang lalu, yang menghasilkan sebagian besar populasi manusia di Eurasia. Diperkirakan bahwa kelompok kedua ini mungkin memiliki teknologi peralatan "berburu hewan besar" yang lebih canggih dan tidak terlalu bergantung pada sumber makanan di pesisir pantai dibandingkan kelompok aslinya. Banyak bukti bahwa ekspansi kelompok pertama dapat dihancurkan oleh naiknya permukaan air laut pada akhir setiap glasial maksimum. Model penyebaran ganda bertentangan dengan penelitian yang menunjukkan bahwa populasi Eurasia serta populasi Asia Tenggara dan Oseania semuanya merupakan keturunan dari garis keturunan DNA mitokondria L3 yang sama, yang mendukung migrasi tunggal keluar dari Afrika yang memunculkan semua populasi non-Afrika.
Atas dasar tanggal awal Badoshan Iran Aurignacian, Oppenheimer mengemukakan bahwa penyebaran kedua ini mungkin terjadi dengan periode pluvial sekitar 50.000 tahun sebelum sekarang, dengan manusia modern yang memiliki budaya berburu hewan besar menyebar ke Pegunungan Zagros, membawa genom manusia modern dari Oman, di seluruh Teluk Persia, ke utara ke Armenia dan Anatolia, dengan sebuah varian berjalan ke selatan ke Israel dan ke Kirenaika.
Bukti genetik terkini menunjukkan bahwa semua populasi modern non-Afrika, termasuk yang ada di Eurasia dan Oseania, adalah keturunan dari gelombang tunggal yang meninggalkan Afrika antara 65.000 dan 50.000 tahun yang lalu.
Penggunaan alat-alat




Penggunaan alat telah diinterpretasikan sebagai tanda kecerdasan, dan ditenggarai bahwa penggunaan alat mungkin telah menstimulasi aspek-aspek tertentu dalam evolusi manusia, terutama perluasan otak manusia yang berkesinambungan. Paleontologi belum dapat menjelaskan secara terperinci mengenai perluasan organ ini dalam rentang jutaan tahun, meskipun peran organ ini yang membutuhkan konsumsi energi yang sangat besar. Otak manusia modern mengkonsumsi sekitar 13 watt (260 kilokalori) per hari, seperlima dari konsumsi daya istirahat tubuh. Peningkatan penggunaan alat memungkinkan perburuan terhadap hewan-hewan dan tumbuhan yang memiliki konten energi yang lebih tinggi. Para peneliti berpendapat bahwa hominin purba berada di bawah tekanan evolusi untuk meningkatkan kapasitas mereka dalam menciptakan dan menggunakan alat.
Kapan tepatnya manusia purba mulai menggunakan alat adalah hal yang sulit untuk ditentukan, karena semakin primitif alat tersebut (misalnya, batu-batu bermata tajam), semakin sulit pula untuk menentukan apakah alat tersebut adalah benda alam atau artefak manusia. Ada beberapa bukti bahwa manusia purba australopithecus (4 juta tahun yang lalu) mungkin telah menggunakan patahan tulang sebagai alat, namun hal ini masih diperdebatkan.
Ada banyak spesies yang membuat dan menggunakan alat, namun genus manusialah yang mendominasi area pembuatan dan penggunaan alat yang lebih kompleks. Alat tertua yang diketahui adalah batu yang diserpih yang ditemkan di Turkana Barat, Kenya, yang berasal dari 3,3 juta tahun yang lalu. Alat-alat batu tertua berikutnya berasal dari Gona, Ethiopia, dan dianggap sebagai awal dari teknologi Oldowan. Alat-alat tersebut berasal dari sekitar 2,6 juta tahun yang lalu. Sebuah fosil Homo ditemukan di dekat beberapa peralatan Oldowan, dan usianya tercatat 2,3 juta tahun, yang menunjukkan bahwa kemungkinan spesies Homo memang membuat dan menggunakan alat-alat ini. Ini adalah sebuah kemungkinan, namun belum menjadi bukti yang kuat. Proses styloid metakarpal ketiga memungkinkan tulang tangan mengunci ke dalam tulang pergelangan tangan, sehingga memungkinkan tekanan yang lebih besar untuk diterapkan pada pergelangan tangan dan tangan dari ibu jari dan jari-jari yang menggenggam. Hal ini memungkinkan manusia memiliki ketangkasan dan kekuatan untuk membuat dan menggunakan alat yang kompleks. Fitur anatomi yang unik ini memisahkan manusia dari kera dan primata bukan manusia lainnya, dan tidak terlihat pada fosil manusia yang berusia lebih dari 1,8 juta tahun.
Bernard Wood mengatakan bahwa Paranthropus hidup berdampingan dengan spesies Homo awal di daerah "Kompleks Industri Oldowan" dalam rentang waktu yang kurang lebih sama. Meskipun tidak ada bukti langsung yang mengidentifikasi Paranthropus sebagai pembuat alat, anatomi mereka memberikan bukti tidak langsung tentang kemampuan mereka di bidang ini. Sebagian besar ahli paleoantropologi setuju bahwa spesies Homo awal memang bertanggung jawab atas sebagian besar peralatan Oldowan yang ditemukan. Mereka berpendapat bahwa ketika sebagian besar peralatan Oldowan ditemukan bersamaan dengan fosil manusia, Homo selalu ada, tapi Paranthropus tidak.
Pada tahun 1994, Randall Susman menggunakan anatomi ibu jari yang berlawanan sebagai dasar argumennya bahwa spesies Homo dan Paranthropus adalah pembuat perkakas. Dia membandingkan tulang dan otot ibu jari manusia dan simpanse, dan menemukan bahwa manusia memiliki 3 otot yang tidak dimiliki simpanse. Manusia juga memiliki metakarpal yang lebih tebal dengan kepala yang lebih lebar, memungkinkan genggaman yang lebih tepat daripada yang bisa dilakukan oleh tangan simpanse. Susman berpendapat bahwa anatomi modern ibu jari manusia yang berlawanan merupakan respons evolusioner terhadap kebutuhan yang terkait dengan pembuatan dan penanganan alat, dan bahwa kedua spesies tersebut memanglah pembuat alat.
Transisi ke perilaku modern
Para antropolog menggambarkan perilaku manusia modern mencakup ciri-ciri budaya dan perilaku seperti spesialisasi alat, penggunaan perhiasan dan gambar (seperti gambar gua), penataan ruang hidup, ritual (seperti hadiah kuburan), teknik berburu khusus, eksplorasi wilayah geografis yang kurang ramah, dan jaringan perdagangan barter, serta ciri-ciri yang lebih umum seperti bahasa dan pemikiran simbolis yang kompleks. Perdebatan terus berlanjut mengenai apakah sebuah "revolusi" menyebabkan manusia modern ("ledakan besar kesadaran manusia"), atau apakah evolusi itu lebih bertahap.
Sampai sekitar 50.000-40.000 tahun yang lalu, penggunaan alat-alat batu tampaknya berkembang secara bertahap. Setiap fase (H. habilis, H. ergaster, H. neanderthalensis) menandai sebuah teknologi baru, diikuti dengan perkembangan yang sangat lambat hingga fase berikutnya. Saat ini para ahli paleoantropologi memperdebatkan apakah para spesies Homo tersebut memiliki beberapa atau banyak perilaku manusia modern. Mereka tampaknya memiliki budaya konservatif, mempertahankan teknologi dan pola makan yang sama dalam waktu yang sangat lama.
Sekitar tahun 50.000 sebelum sekarang, budaya manusia mulai berevolusi dengan lebih cepat. Transisi menuju modernitas perilaku telah dicirikan oleh beberapa peneliti sebagai "Lompatan Besar ke Depan", atau sebagai "Revolusi Palaeolitik Akhir", karena kemunculan mendadak dalam catatan arkeologi tanda-tanda khas perilaku modern dan perburuan hewan buruan. Bukti perilaku modernitas yang jauh lebih awal juga ditemukan di Afrika, dengan bukti-bukti yang lebih tua berupa gambar-gambar abstrak, strategi bertahan hidup yang lebih luas, peralatan dan senjata yang lebih canggih, dan perilaku "modern" lainnya, dan banyak peneliti baru-baru ini berargumen bahwa transisi menuju modernitas terjadi lebih cepat daripada yang diyakini sebelumnya. Beberapa peneliti lain menganggap transisi tersebut lebih bertahap, dengan menekankan bahwa beberapa fitur telah muncul di antara Homo sapiens Afrika purba 300.000-200.000 tahun yang lalu. Bukti terbaru menunjukkan bahwa penduduk Aborigin Australia terpisah dari penduduk Afrika 75.000 tahun yang lalu, dan bahwa mereka melakukan perjalanan laut sejauh 160 km 60.000 tahun yang lalu, yang mungkin mengurangi signifikansi Revolusi Paleolitik Akhir.
Manusia modern mulai menguburkan mayat mereka, membuat pakaian dari kulit binatang, berburu dengan teknik yang lebih canggih (seperti menggunakan lubang jebakan atau menggiring binatang agar jatuh dari tebing), dan melukis di gua. Seiring dengan kemajuan budaya manusia, populasi yang berbeda berinovasi dengan teknologi yang ada: artefak seperti kail ikan, kancing, dan jarum tulang menunjukkan tanda-tanda variasi budaya, yang belum pernah terlihat sebelum tahun 50.000 SM. Biasanya, populasi H. neanderthalensis yang lebih tua tidak memiliki variasi dalam teknologi mereka, meskipun kumpulan Chatelperronian telah ditemukan sebagai tiruan oleh Neanderthal atas teknologi Aurignasia-nya H. sapiens.
Evolusi manusia terkini dan yang sedang berlangsung
Populasi manusia modern secara anatomis senantiasa berevolusi, karena dipengaruhi oleh seleksi alam dan pergeseran genetik. Meskipun tekanan seleksi pada beberapa sifat, seperti ketahanan terhadap cacar, telah berkurang di era modern, manusia masih mengalami seleksi alam untuk banyak sifat lainnya. Beberapa di antaranya disebabkan oleh tekanan lingkungan tertentu, sementara yang lain terkait dengan perubahan gaya hidup sejak perkembangan pertanian (10.000 tahun yang lalu), urbanisasi (5.000), dan industrialisasi (250 tahun yang lalu). Telah diperdebatkan bahwa evolusi manusia telah dipercepat sejak perkembangan pertanian 10.000 tahun yang lalu dan perkembangan peradaban sekitar 5.000 tahun yang lalu, yang mengakibatkan, perbedaan genetik yang substansial antara beragam populasi manusia saat ini, dan penelitian yang lebih baru mengindikasikan bahwa untuk beberapa sifat, perkembangan dan inovasi budaya manusia telah mendorong bentuk seleksi baru yang hidup berdampingan bersama, dan dalam beberapa kasus sebagian besar telah menggantikan seleksi alam.
Yang paling mencolok adalah variasi dalam karakteristik superfisial, seperti rambut bertekstur Afro, atau evolusi kulit putih dan rambut pirang pada beberapa populasi, yang disebabkan oleh perbedaan iklim. Tekanan selektif yang sangat kuat telah menghasilkan adaptasi di dataran tinggi pada manusia, dengan adaptasi yang berbeda-beda pada tiap-tiap populasi yang terisolasi. Studi tentang dasar genetik menunjukkan bahwa beberapa mengembangkan sifat-sifat yang baru belakangan ini, dengan orang Tibet berevolusi lebih dari 3.000 tahun untuk memiliki proporsi alel EPAS1 yang tinggi yang adaptif terhadap dataran tinggi.
Evolusi lainnya terkait dengan penyakit endemik: keberadaan malaria menyeleksi sifat sel sabit (bentuk heterozigot gen sel sabit), sementara jika tidak ada malaria, efek kesehatan anemia sel sabit akan menyeleksi melawan sifat ini. Sebagai contoh lain, populasi yang berisiko terkena penyakit kuru yang menyebabkan kelemahan parah memiliki representasi berlebihan yang signifikan pada varian imun dari gen protein prion G127V dibandingkan dengan alel non-imun. Munculnya frekuensi varian genetik ini disebabkan oleh ketahanan pada orang yang imun dari penyakit tersebut. Tren lainnya yang masih belum dapat dijelaskan dan menjadi subjek penelitian yang sedang berlangsung di bidang baru kedokteran evolusi: sindrom ovarium polikistik mengurangi kesuburan sehingga diperkirakan akan mengalami seleksi negatif yang sangat kuat, namun kesamaan relatifnya dalam populasi manusia menunjukkan adanya tekanan seleksi yang berlawanan. Identitas dari tekanan tersebut masih menjadi bahan perdebatan.
Evolusi manusia baru-baru ini yang terkait dengan pertanian mencakup resistensi genetik terhadap penyakit menular yang telah muncul pada populasi manusia yang melintasi penghalang spesies dari hewan peliharaan, serta perubahan metabolisme dikarenakan perubahan pola makan, seperti persistensi laktase.
Bukti
Bukti-bukti yang mendasari catatan ilmiah tentang evolusi manusia berasal dari banyak bidang ilmu pengetahuan alam. Sumber utama pengetahuan tentang proses evolusi secara tradisional adalah catatan fosil, tetapi sejak perkembangan genetika yang dimulai pada tahun 1970-an, analisis DNA telah menempati tempat yang sama pentingnya. Studi tentang ontogeni, filogeni dan terutama biologi perkembangan evolusioner dari vertebrata dan invertebrata menawarkan wawasan yang cukup besar ke dalam evolusi semua kehidupan, termasuk bagaimana manusia berevolusi. Studi khusus tentang asal-usul dan kehidupan manusia adalah antropologi, khususnya paleoantropologi yang berfokus pada studi prasejarah manusia.
Bukti dari genetika

Kerabat terdekat manusia yang masih hidup adalah bonobo dan simpanse (keduanya genus Pan) dan gorila (genus Gorilla). Dengan pengurutan genom manusia dan simpanse, hingga 2012 diperkirakan kemiripan antara sekuens DNA mereka berkisar antara 95% dan 99%. Dengan menggunakan teknik yang disebut jam molekuler yang memperkirakan waktu yang diperlukan untuk jumlah mutasi divergen untuk mengakumulasi di antara dua garis keturunan, perkiraan tanggal pemisahan antar garis keturunan dapat dihitung.
Owa (famili Hylobatidae) dan kemudian orang utan (genus Pongo) adalah kelompok pertama yang memisahkan diri dari garis yang mengarah ke hominin, termasuk manusia-diikuti oleh gorila (genus Gorilla), dan, akhirnya, oleh simpanse (genus Pan). Tanggal pemisahan antara garis keturunan hominin dan simpanse ditempatkan oleh beberapa kalangan antara 4 hingga 8 juta tahun silam, yakni selama Miosen Akhir. Akan tetapi, spesiasi tampaknya telah terjadi secara berlarut-larut. Pemisahan awal terjadi antara 7 hingga 13 juta tahun silam, tetapi hibridisasi yang terus berlangsung mengaburkan pemisahan dan menunda pemisahan total selama beberapa juta tahun. Patterson (2006) memperkirakan pemisahan akhir pada 5 hingga 6 juta tahun silam.
Bukti genetik juga telah digunakan untuk membandingkan spesies di dalam genus Homo, menyelidiki aliran gen antara manusia modern awal dan Neanderthal, dan untuk lebih dapat memahami pola migrasi manusia awal dan waktu pemisahannya. Dengan membandingkan bagian-bagian genom yang tidak berada di bawah seleksi alam dan oleh karena itu mengakumulasi mutasi pada tingkat yang cukup konstan, adalah mungkin untuk merekonstruksi pohon genetik yang mencakup seluruh spesies manusia sejak nenek moyang bersama terakhir.
Setiap kali mutasi tertentu (polimorfisme nukleotida tunggal) muncul pada seorang individu dan diwariskan kepada keturunannya, sebuah haplogroup terbentuk mencakup seluruh keturunan individu tersebut yang akan membawa mutasinya. Dengan membandingkan DNA mitokondria yang hanya diwarisi dari ibu, para ahli genetika telah menyimpulkan bahwa nenek moyang bersama wanita terakhir yang penanda genetik-nya ditemukan pada semua manusia modern, yang disebut Hawa mitokondria, pastinya hidup sekitar 200.000 tahun yang lalu.
Genetika
Genetika evolusioner manusia mempelajari perbedaan genom manusia di antara tiap-tiap individu, evolusi masa lalu yang memunculkan genom-genom tersebut, dan efeknya pada saat ini. Perbedaan di antara genom memiliki implikasi dan aplikasi antropologis, medis dan forensik. Data genetik dapat memberikan wawasan penting tentang evolusi manusia.
Bukti dari rekaman fosil


Tidak terdapat banyak bukti fosil untuk pemisahan garis keturunan gorila, simpanse, dan hominin. Fosil-fosil paling tua yang telah diajukan sebagai anggota garis keturunan hominin adalah Sahelanthropus tchadensis yang berasal dari 7 juta tahun silam, Orrorin tugenensis yang berasal dari 5,7 juta tahun silam, dan Ardipithecus kadabba yang berasal dari 5,6 juta tahun silam. Masing-masing telah diperdebatkan sebagai nenek moyang bipedal dari hominin yang muncul kemudian, tetapi, dalam tiap-tiap kasus, masing-masing klaim diperdebatkan. Terdapat pula kemungkinan bahwa satu atau lebih spesies ini merupakan nenek moyang dari cabang lain kera Afrika, atau bahwa mereka mewakili nenek moyang bersama antara hominin dan kera lainnya.
Pertanyaan tentang hubungan antara spesies fosil kuno ini dengan garis keturunan hominin masih harus dipecahkan. Dari spesies-spesies awal ini, australopithecine muncul sekitar 4 juta tahun silam dan berpisah menjadi cabang-cabang yang berpostur tegap (juga disebut Paranthropus) dan ramping, yang salah satunya (kemungkinan A. garhi) mungkin adalah nenek moyang dari genus Homo. Spesies australopithecine yang paling baik terwakili dalam catatan fosil adalah Australopithecus afarensis dengan lebih dari 100 individu fosil yang ditemukan mulai dari Etiopia Utara (seperti "Lucy" yang terkenal), hingga Kenya, dan Afrika Selatan. Fosil australopithecines yang tegap seperti Au. robustus (atau Paranthropus robustus) dan Au./ P. boisei sangat melimpah di Afrika Selatan di situs-situs seperti Kromdraai dan Swartkrans, dan di sekitar Danau Turkana di Kenya.
Anggota paling awal dari genus Homo adalah Homo habilis yang berevolusi sekitar 2,8 juta tahun silam. Homo habilis adalah spesies pertama yang memiliki bukti positif tentang penggunaan peralatan batu. Mereka mengembangkan teknologi litik Oldowan, dinamai sesuai dengan nama Ngarai Olduvai di mana spesimennya pertama kali ditemukan. Beberapa ilmuwan menganggap Homo rudolfensis, kelompok fosil bertubuh lebih besar dengan morfologi yang mirip dengan fosil H. habilis asli, sebagai spesies yang terpisah, sementara yang lain menganggap mereka sebagai bagian dari H. habilis - hanya mewakili variasi intraspesies, atau bahkan mungkin dimorfisme seksual. Otak hominin purba ini berukuran hampir sama dengan simpanse, dan adaptasi utama mereka adalah bipedalisme sebagai penyesuaian diri terhadap kehidupan terestrial.
Selama jutaan tahun berikutnya, proses ensefalisasi dimulai dan, pada saat kedatangan (sekitar 1,9 juta tahun silam) Homo erectus dalam catatan fosil, kapasitas tempurung kepalanya telah berlipat ganda. Homo erectus adalah hominin pertama yang beremigrasi dari Afrika. Dan, dari 1,8 hingga 1,3 juta tahun silam, spesies ini menyebar melalui Afrika, Asia, dan Eropa. Satu populasi dari H. erectus, yang juga terkadang diklasifikasikan sebagai spesies terpisah Homo ergaster, tetap berada di Afrika dan berevolusi menjadi Homo sapiens. Dipercaya bahwa kedua spesies ini, H. erectus dan H. ergaster, adalah yang pertama kali menggunakan api dan peralatan yang kompleks. Di Eurasia H. erectus berevolusi menjadi spesies seperti H. antecessor, H. heidelbergensis dan H. neanderthalensis. Fosil-fosil paling tua dari manusia modern secara anatomis berasal dari Paleolitikum Tengah, sekitar 300-200.000 tahun yang lalu seperti sisa-sisa Omo dan Herto di Ethiopia, sisa-sisa Jebel Irhoud di Maroko, dan sisa-sisa Florisbad di Afrika Selatan; fosil-fosil selanjutnya dari gua Es Skhul di Israel dan Eropa Selatan dimulai sekitar 90.000 tahun yang lalu (0,09 juta tahun silam).
Seiring dengan penyebaran manusia modern dari Afrika, mereka bertemu dengan hominin lain seperti Homo neanderthalensis dan Denisovan, yang mungkin telah berevolusi dari populasi Homo erectus yang telah meninggalkan Afrika sekitar 2 juta tahun silam. Sifat interaksi antara manusia purba dan spesies saudari mereka ini telah menjadi sumber kontroversi berkepanjangan, pertanyaannya adalah apakah manusia menggantikan spesies-spesies yang lebih awal ini atau apakah mereka sebenarnya cukup mirip hingga dapat melakukan kawin silang, yang mana dalam hal ini populasi-populasi yang lebih awal ini mungkin telah menyumbangkan materi genetik kepada manusia modern.
Migrasi keluar dari Afrika ini diperkirakan dimulai sekitar 70-50.000 tahun BP dan manusia modern kemudian menyebar secara global, menggantikan hominin sebelumnya baik melalui kompetisi atau hibridisasi. Mereka mulai mendiami Eurasia dan Oseania pada 40.000 tahun BP, dan Amerika setidaknya pada 14.500 tahun BP.
Perkawinan antar spesies
Hipotesis adanya kawin silang, juga dikenal sebagai hibridisasi, pencampuran atau teori asal-usul hibrida, telah didiskusikan sejak ditemukannya sisa-sisa Neanderthal pada abad ke-19. Pandangan yang mengatakan bahwa evolusi manusia itu linier mulai ditinggalkan pada tahun 1970-an karena ditemukannya berbagai spesies manusia yang berbeda, yang membuat konsep linier semakin tidak memungkinkan. Pada abad ke-21 dengan munculnya teknik biologi molekuler dan komputerisasi, pengurutan seluruh genom Neanderthal dan genom manusia dilakukan, yang mengkonfirmasi adanya perkawinan silang antara spesies-spesies manusia yang berbeda. Pada tahun 2010, bukti berdasarkan biologi molekuler diterbitkan, mengungkapkan contoh-contoh yang gamblang dari perkawinan silang antara manusia purba dan modern selama Paleolitik Tengah dan awal Paleolitik Atas. Telah ditunjukkan bahwa perkawinan silang terjadi dalam beberapa peristiwa independen yang mencakup Neanderthal dan Denisovan, serta beberapa hominin yang tidak teridentifikasi. Saat ini, sekitar 2% DNA dari semua populasi non-Afrika (termasuk Eropa, Asia, dan Oseania) adalah Neanderthal, dengan sejumlah jejak DNA warisan Denisovan. Juga, 4-6% genetika orang Melanesia modern adalah Denisovan. Perbandingan genom manusia dengan genom Neandertal, Denisovan, dan kera dapat membantu mengidentifikasi fitur-fitur yang membedakan manusia modern dari spesies hominin lainnya. Dalam studi genomik komparatif 2016, tim peneliti Sekolah Medis Harvard/UCLA membuat peta dunia tentang distribusi dan membuat sejumlah prediksi tentang di mana gen Denisovan dan Neanderthal mungkin berdampak pada biologi manusia modern.
Sebagai contoh, studi komparatif pada pertengahan tahun 2010-an menemukan beberapa sifat yang berkaitan dengan fenotip metabolik, perkembangan, neurologis, dan imunologis, yang dikembangkan oleh manusia purba ke lingkungan Eropa dan Asia, dan diwariskan kepada manusia modern melalui pencampuran dengan hominin lokal.
Meskipun narasi evolusi manusia sering kali diperdebatkan, beberapa penemuan sejak tahun 2010 menunjukkan bahwa evolusi manusia seharusnya tidak dilihat sebagai perkembangan linear atau bercabang yang sederhana, tetapi campuran antara spesies yang terkait. Faktanya, penelitian genomik telah menunjukkan bahwa hibridisasi antara garis keturunan yang berpisah secara substansial adalah sebuah kelaziman, bukan pengecualian, dalam evolusi manusia. Lebih jauh lagi, dikatakan bahwa hibridisasi adalah daya kreatif yang penting dalam kemunculan manusia modern.