

| Bagian dari seri |
| Paleontologi |
|---|
 |
| Kategori |
| Bagian dari seri |
| Paleontologi |
|---|
|
| Kategori |
Fosil peralihan atau bentuk peralihan atau fosil transisi adalah sisa-sisa bentuk kehidupan yang menjadi fosil dan menunjukkan ciri-ciri dari dua kelompok taksonomi yang berbeda. Fosil peralihan merupakan fosil organisme yang berada di dekat titik percabangan ketika garis keturunan individual utama (klad) terpisah. Fosil peralihan memiliki ciri-ciri umum dari organisme pada kedua bagian pemisahannya, tetapi karena kurang lengkapnya rekaman fosil, biasanya tidak ada cara untuk mengetahui secara pasti seberapa dekatnya suatu fosil peralihan dengan titik pasti pemisahannya.
Fosil peralihan berguna sebagai pengingat bahwa pembagian taksonomi merupakan buatan manusia yang telah diberlakukan dalam kontinum variasi. Banyak bukti mengenai keberadaan fosil peralihan, termasuk di antaranya adalah fosil peralihan antara manusia dan primata lainnya, antara tetrapoda dan ikan, dan antara burung dan dinosaurus. Istilah "mata rantai yang hilang" sering sekali digunakan dalam tulisan-tulisan populer mengenai evolusi manusia untuk merujuk kepada celah yang dianggap ada dalam catatan evolusi hominid. Istilah ini biasanya digunakan untuk merujuk kepada setiap temuan fosil peralihan terbaru. Akan tetapi, para ilmuwan tidak menggunakan istilah "mata rantai yang hilang" karena istilah ini tidak akurat dan menyesatkan.
Evolusi
| 1850 |

|
| 1900 |

|
| 1950 |

|
| 2002 |
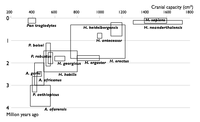
|
| Diagram-diagram ini menunjukkan spesies-spesies Homininae yang diketahui pada tahun-tahun tersebut. Tiap spesies ditunjukkan dalam kotak yang memperlihatkan jangkauan kapasitas kranial dari tiap spesimen spesies yang bersangkutan, dan jangkauan waktu kehidupan tiap spesies di bumi. Rangkaian diagram ini menunjukkan bagaimana "mata rantai yang hilang" atau celah antara tiap spesies dalam rekaman fosil menjadi terisi seiring ditemukannya lebih banyak fosil. | |
Pada tahun 1859, ketika buku On the Origin of Species karya Charles Darwin pertama kali diterbitkan, rekaman fosil yang ada sangatlah minim, dan Darwin mengungkapkan bahwa kurangnya fosil peralihan merupakan "tantangan yang paling jelas dan paling berbahaya terhadap teori saya", tetapi dia menjelaskannya dengan menghubungkannya dengan ketidaksempurnaan yang ekstrem dari rekaman geologis. Dia mengamati koleksi yang terbatas pada waktu itu, tetapi menggambarkan informasi yang tersedia sebagai pola penunjuk yang mengikuti teorinya mengenai keturunan dengan perubahan melalui seleksi alam. Dan memang, Archaeopteryx ditemukan hanya dua tahun kemudian, tepatnya pada tahun 1861, dan menunjukkan bentuk peralihan klasik antara dinosaurus dan burung. Sejak saat itu, banyak sekali fosil peralihan yang telah ditemukan dan kini dianggap bahwa ada banyak bukti mengenai bagaimana semua kelas vertebrata saling berkaitan, banyak di antaranya dalam bentuk fosil peralihan.
Contoh
Rekonstruksi evolusi kuda dan kerabat-kerabatnya dikumpulkan oleh Othniel Charles Marsh dari fosil-fosil yang ditemukan yang berhasil membentuk suatu garis keturunan tunggal yang berkembang secara konsisten dengan banyak jenis "peralihan", dan kadang disebut sebagai pohon keluarga. Akan tetapi, kladistika modern memberikan gambaran berbeda yang mirip semak bercabang banyak, dengan banyak perubahan dan banyak pula jalur buntu. Spesimen lainnya yang sering disebut sebagai bentuk peralihan meliputi "paus berjalan" Ambulocetus, ikan bersirip cuping yang belum lama ini ditemukan Tiktaalik serta beragam hominid yang dipercaya sebagai manusia proto.
Nenek moyang tumbuhan berbiji dari periode Devon pertengahan dari Belgia telah teridentifikasi muncul 20 juta tahun lebih dahulu daripada tumbuhan berbiji pertama. Runcaria, yang berbentuk kecil dan bersimetri radial, adalah megasporangium yang berintegumen dan dikelilingi oleh kupule. Megasporangium itu menghasilkan distal tertutup yang memanjang dan menonjol di atas integumen bercuping banyak. Diduga bahwa perpanjangan itu berperan dalam penyerbukan anemofili. Runcaria memberikan pemahaman baru kepada pengenalan ciri-ciri yang berujung pada biji. Runcaria memiliki cemua ciri tumbuhan berbiji kecuali kulit biji yang keras serta sistem untuk memandu serbuk sari menuju ke biji.
Keterbatasan rekaman fosil
Tidak semua bentuk peralihan muncul dalam rekaman fosil karena rekaman fosil belumlah lengkap. Makhluk hidup jarang tersimpan menjadi fosil karena proses menjadi fosil hanya dapat terjadi pada keadaan yang terbaik dan hanya bagian-bagian fosil semacam itu yang sudah ditemukan. Paleontolog Donald Prothero mengamati bahwa hal ini ditunjukkan oleh fakta bahwa jumlah keseluruhan spesies dari semua jenis yang diketahui melalui rekaman fosil adalah kurang dari lima persen dari jumlah spesies hidup yang diketahui, yang mengindikasikan bahwa jumlah spesies yang diketahui melalui fosil pastilah kurang dari satu persen dari seluruh spesies yang pernah hidup di bumi.
Rekaman fosil sangat tidak berimbang, dan dengan beberapa pengecualian, rekaman fosil yang ada sekarang kebanyakan merupakan fosil yang berasal dari organisme yang memiliki bagian tubuh yang keras, sementara sebagian besar organisme bertubuh lunak hanya meninggalkan rekaman fosil yang amat sangat sedikit. Kelompok yang dianggap memiliki rekaman fosil yang bagus, meliputi sejumlah fosil peralihan antara kelompok hewan tradisional, antara lain vertebrata, echinodermata, brachiopoda serta beberapa kelompok arthropoda.

Taksonomi evolusioner dan kladistika
Dalam taksonomi evolusioner, yang merupakan bentuk taksonomi paling umum selama sebagian besar abad ke-20 dan masih digunakan dalam buku-buku teks dasar, taksa yang didasarkan pada kemiripan morfologis sering kali digambarkan sebagai "gelembung-gelembung" yang saling bercabang namun terpisah satu sama lain, membentuk pohon evolusi. Bentuk peralihan dilihat sebagai penyambung di antara beragam kelompok dalam hal anatomi, dan dengan demikian ditaruh di perbatasan di antara gelembung-gelembung tersebut.
Dengan berdirinya metode kladistika, hubungannya kini secara ketat ditunjukkan dalam apa yang disebut kladogram, yang menunjukkan percabangan garis keturunan dalam evolusi. Kelompok-kelompok berbeda yang disebut 'natural' atau 'monofiletik' membentuk unit bersarang yang tidak saling tumpang tindih. Dengan demikian dalam kladistika tidak ada lagi peralihan antara kelompok-kelompk yang telah ada, melainkan diferensiasi yang berlangsung di dalam kelompok, yang dilambangkan sebagai cabang dalam kladogram. Dalam konteks ini, organisme peralihan dapat dikonseptualisasikan sebagai yang perwakilan dari contoh-contoh awal pada cabang-cabang yang berbeda pada kladogram, terletak di antara titik cabang tertentu dan "kelompok mahkota", yang merupakan kelompok yang paling diturunkan, yang diletakkan di ujung suatu garis keturunan.
Peralihan vs moyang
Ada suatu kesalahpahaman yang disebabkan oleh suatu konsep bahwa bentuk peralihan antara dua kelompok taksonomi berbeda pastilah merupakan moyang secara langsung bagi satu atau dua kelompok keturunannya. Ini ditambah lagi oleh fakta bahwa tujuan taksonomi evolusioner adalah untuk berusaha mengidentifikasi taksa yang merupakan moyang bagi taksa lainnya. Akan tetapi, hampir tidak mungkin untuk dapat yakin bahwa setiap bentuk yang ditunjukkan dalam rekaman fosil merupakan leluhur langsung dari makhluk lainnya. Pada kenyataannya, karena evolusi merupakan suatu proses yang bercabang yang menghasilkan pola rumit mirip semak yang menunjukkan spesies yang saling berkaitan dan bukannya suatu proses linear yang menghasilkan perkembangan mirip tangga, dan karena tidak lengkapnya rekaman fosil, agak tidak mungkin jika bentuk apapun yang ditunjukkan dalam rekaman fosil merupakan leluhur langsung bagi yang lainnya. Kladistika sangat tidak menekankan konsep mengenai satu kelompok taksonomi sebagai nenek moyang bagi yang lainnya, malah menekankan konsep mengidentifikasi taksa bersaudara yang saling memiliki leluhur yang sama secara lebih terkini daripada dengan kelompok lainnya. Ada beberapa kasus pengecualian, misalnya beberapa fosil mikro plankton laut, yang mana rekaman fosilnya cukup lengkap untuk menunjukkan secara cukup yakin bahwa fosil-fosil tertentu mewakili populasi yang sebenarnya merupakan nenek moyang bagi populasi setelahnya yang merupakan sepesies yang berbeda, tetapi secara umum fosil peralihan dianggap memiliki ciri-ciri yang menunjukkan ciri anatomi peralihan dari leluhur bersama yang sebenarnya bagi beberapa taksa dan bukannya leluhur itu sendiri.
Mata rantai yang hilang
Istilah "mata rantai yang hilang" merujuk kepada konsep statis praevolusi tentang rantai keberadaan, sebuah gagasan deisme bahwa semua keberadaan terhubung, mulai dari tanah, melalui kerajaan makhluk hidup hingga malaikat dan akhirnya hingga tuhan. Gagasan bahwa semua makhluk hidup terhubung melalui semacam proses transmutasi telah muncul sebelum teori evolusi Darwin. Jean-Baptiste Lamarck berpendapat bahwa kehidupan dihasilkan dalam bentuk makjluk paling sederhana secara konstan, dan kemudian berjuang menuju kesempurnaan dan kerumitan (yaitu manusia) melalui serangkaian bentuk lebih rendah. Dalam pandangannya, hewan lebih rendah hanyalah pendatang baru dalam panggung evolusi.
Pada kenyataannya, semakin lama semakin banyak fosil peralihan yang ditemukan. Ini semakin menambah pengetahuan mengenai proses peralihan evolusi itu sendiri, dan semakin banyak pula penemuan fosil peralihan yang dulunya dianggap sebagai "mata rantai yang hilang". Ini menjadikan istilah "mata rantai yang hilang" sebagai istilah yang tidak akurat.
Istlah "mata rantai yang hilang" digunakan oleh Charles Lyell dalam cara yang cukup berbeda dalam karyanya, Elements of Geology, pada tahun 1851, tetapi menjadi terkenal dengan maknanya yang sekarang berkat kemunculan istilah ini dalam karya Lyell lainnya yang berjudul Geological Evidences of the Antiquity of Man pada tahun 1863 pada halaman xi. Pada masa itu para geolog mengabaikan penafsiran injil secara harfiah dan secara umum dipercaya bahwa akhir periode glasial terakhir menandai kemunculan pertama umat manusia, suatu pendapat yang dikemukakan oleh Lyell dalam karyanya Element of Geology. Sementara itu Geological Evidences of the Antiquity of Man mendasarkan pada temuan-temuan baru untuk menempatkan asal usul manusia jauh lebih tua pada masa lalu geologis yang lebih purba. Tulisan Lyell, yang jelas dan hidup, memicu imajinasi publik, dan menginspirasi Jules Verne dalam karyanya Journey to the Center of the Earth, serta menginspirasi juga edisi kedua dari La Terre avant le déluge karya Louis Figuier pada tahun 1867, yang menyertakan ilustrasi dramatis mengenai pria dan wanita purba yang mengenakan kulit binatang dan membawa-bawa kapak batu, menggantikan ilustrasi Taman Eden, yang ditampilkan pada edisi pertamanya pada tahun 1863.
Gagasan mengenai "mata rantai yang hilang" antara manusia dan hewan "yang lebih rendah" tetap tersimpan dalam imajinasi publik. Konsep ini menjadi semakin terkenal dengan ditemukannya Australopithecus africanus (Bocah Taung), Australopithecus sediba,Homo erectus (Manusia Peking, Manusia Jawa, Bocah Turkana), dan fosil-fosil Hominina lainnya.
Kesalahpahaman


Para pendukung kreasionisme sangat sering membuat klaim mengenai keberadaan atau implikasi dari fosil peralihan. Banyak dari klaim yang dibuat oleh para kreasonis itu tidak dianggap benar oleh kalangan paleontolog, dan dalam beberapa kasus klaim dari para kreasionis adalah menyesatkan. Beberapa klaim tersebut adalah:
- 'Tidak ada fosil peralihan.' Ini adalah klaim yang dinyatakan oleh kelompok-kelompok semacam Answers in Genesis dan Institute for Creation Research. Klaim semacam ini mungkin didasarkan pada kesalahpahaman konsep mengenai apa yang dianggap sebagai ciri-ciri peralihan. Klaim ini juga dianggap sebagai siasat yang digunakan oleh para kreasionis untuk memutarbailkkan atau mendiskreditkan teori evolusi dan disebut sebagai "kebohongan kreasionis yang paling disukai". Sementara itu beberapa kreasionis memperdebatkan kurangnya bentuk peralihan.
- 'Tidak ada fosil yang ditemukan memiliki organ tubuh yang tidak berfungsi secara penuh.' Pada kenyataanya, banyak contoh organ vestigial (tidak memiliki fungsi penuh) pada makhluk hidup, misalnya kaki paus, sayap burung yang tak dapat terbang, panggul dan paru-paru ular, serta banyak sekali organ dalam tubuh manusia, di antaranya adalah tulang ekor, plica semilunaris, dan usus buntu.
- Henry M. Morris dan para kreasionis lainnya mengklaim bahwa evolusi memperkirakan perubahan bertahap yang berkelanjutan pada rekaman fosil, dan telah salah memahami rekaman yang hanya ada sebagian sebagai "celah sistematis". Pada kenyataannya, makhluk biologis membutuhkan keadaan yang sangat khusus dan jarang supaya dapat berubah menjadi fosil, akibatnya hanya sebagian kecil dari keseluruhan bentuk kehidupan yang pernah menghuni bumi yang dapat ditemukan dalam bentuk fosil dan tiap temuan fosil hanya merupakan satu kilasan kecil dari proses evolusi itu sendiri. Proses peralihan itu sendiri hanya dapat diilustrasikan dan dikuatkan dengan fosil peralihan, tetapi fosil peralihan tidak akan pernah menunjukkan satu titik pasti yang tepat berada di tengah di antara bentuk terpisah yang berbeda.
Keseimbangan bersela
Teori keseimbangan bersela yang dikembangkan oleh Stephen Jay Gould dan Niles Eldredge dan pertama kali dikemukakan pada tahun 1972 sering kali secara keliru disertakan dalam perbincangan mengenai fosil peralihan. Pada kenyataannya, teori ini hanya membahas peralihan yang terdokumentasi dengan baik dalam satu taksa atau antara taksa yang berkaitan dekat pada periode waktu yang secara geologis pendek. Peralihan ini, biasanya terlacak dalam banyak singkapan geologis, sering kali menunjukkan lompatan-lompatan kecil dalam morfologi antara perpanjangan periode kestabilan morfologi. Untuk menjelaskan lompatan-lompatan ini, Gould dan Eldredge menggambarkan kestabilan genetis pada periode yang relatif lama yang dipisahkan oleh beberapa periode evolusi yang cepat. Gould memberikan komentar berikut mengenai para kreasionis yang menyalahgunakan karyanya untuk membantah keberadaan fosil peralihan:
| “ | Karena kami mengemukakan keseimbangan bersela untuk menjelaskan perkembangan, adalah sangat menyebalkan karena karya kami terus-menerus dikutip oleh para kreasionis—entah secara sengaja ataupun karena kebodohan, aku tidak tahu—untuk menunjukkan bahwa rekaman fosil tidak memiliki bentuk peralihan. Keberselaan terjadi pada tingkatan spesies; kecenderungan arah (pada model tangga) banyak terjadi pada tingkatan peralihan yang lebih tinggi dalam kelompok-kelompok besar. | ” |
| — Stephen Jay Gould | ||
Lihat pula
Catatan kaki
- Browne, E. Janet (2002). Charles Darwin: vol. 2 The Power of Place. London: Jonathan Cape. ISBN 0-7126-6837-3.
- Darwin, Charles (1859). "[[On the Origin of Species]] by Means of Natural Selection, or the Preservation of Favoured Races in the Struggle for Life" (edisi ke-1st). London: John Murray. Konflik URL–wikilink (bantuan)
- Prothero, Donald R. (2007). Evolution: What the Fossils Say and Why it Matters. Columbia University Press. ISBN 978-0-231-13962-5.
Pranala luar
- (Inggris) Transitional vertebrate fossils FAQ, di TalkOrigins Archive.
- (Inggris) A Smooth Fossil Transition: single celled Radiolarian
- (Inggris) Transitional species in insect evolution (Evolusi rayap dari kecoa).