

| |
| Nama | |
|---|---|
| Nama lain
Difosfopiridina nukleotida (DPN+), Koenzim I
| |
| Penanda | |
|
Model 3D (JSmol)
|
|
| 3DMet | {{{3DMet}}} |
| ChEBI | |
| ChemSpider | |
| Nomor EC | |
| KEGG | |
|
PubChem CID
|
|
| Nomor RTECS | {{{value}}} |
| |
| Sifat | |
| C21H27N7O14P2 | |
| Massa molar | 663,43 g/mol |
| Penampilan | bubuk putih |
| Titik lebur | 160 °C |
| Bahaya | |
| Bahaya utama | Tidak berbahaya |
|
Kecuali dinyatakan lain, data di atas berlaku pada temperatur dan tekanan standar (25 °C [77 °F], 100 kPa). | |
| Referensi | |
Nikotinamida adenina dinukleotida, disingkat NAD+, adalah koenzim yang ditemukan di semua sel hidup. Senyawa ini berupa dinukleotida, yakni mengandung dua nukleotida yang dihubungkan melalui gugus fosfat, dengan satu nukleotida mengandung basa adenina dan yang lainnya mengandung nikotinamida.
Dalam metabolisme, NAD+ terlibat dalam reaksi redoks, dengan membawa elektron dari satu reaksi ke reaksi lainnya. Koenzim ini oleh karenanya ditemukan dalam dua bentuk yang berbeda: NAD+ sebagai oksidator, dan NADH sebagai reduktor. NAD+ menerima elektron dari molekul lain dan menjadi tereduksi (NADH), dan begitu pula sebaliknya. Reaksi transfer elektron ini merupakan salah satu fungsi NAD+. Namun ia juga memiliki fungsi lain pada proses seluler lainnya, utamanya adalah sebagai substrat enzim yang menambah maupun mengurangi gugus fungsi pada protein dalam modifikasi pascatranslasional. Karena fungsinya yang penting ini, enzim-enzim yang terlibat dalam metabolisme NAD sering menjadi target pengembangan obat-obatan.
Dalam organisme, NAD+ dapat disintesis secara de novo (dari blok-blok molekul kecil) dari asam amino triptofan ataupun asam aspartat. Selain itu, NAD+ dapat juga diperoleh dari sumber makanan yang mengandung vitamin niasin.
Beberapa NAD diubah menjadi koenzim nikotinamida adenin dinukleotida fosfat (NADP). Struktur kimianya sebagian besar mirip dengan NAD, namun perannya sebagian besar sebagai kofaktor dalam metabolisme anabolik.
Sifat-sifat fisika dan kimia
Nikotinamida adenina dinukleotida, sama seperti senyawa dinukleotida pada umumnya, mengandung nukleotida yang dihubungkan oleh satu pasang gugus fosfat yang menjembatani keduanya. Nukleotida ini tersusun atas cincin ribosa dengan adenina yang melekat pada atom karbon pertama 1' cincin tersebut. Gugus nikotinamida dapat dilekatkan ke dalam dua orientasi pada satu atom karbon anomerik. Karena terdapat dua struktur yang dimungkinkan, senyawa ini berupa diastereomer. Diastereomer β-nikotinamida dari NAD+ adalah bentuk yang ditemukan pada organisme. Kedua nukleotida ini dihubungkan bersama oleh dua gugus fosfat melalui karbon 5'.

Dalam metabolisme, senyawa ini menerima ataupun mendonorkan elektronnya dalam reaksi redoks. Reaksi ini (diringkaskan oleh persamaan di bawah) melibatkan pelepasan dua atom hidrogen dari reaktan (R), dalam bentuk ion hidrida (H−) dan proton (H+). Proton dilepaskan ke dalam larutan, manakala reduktan RH2 dioksidasi dan NAD+ direduksi menjadi NADH melalui transfer hidrida menuju cincin nikotinamida.
- RH2 + NAD+ → NADH + H+ + R
Dari pasangan elektron hidrida, satu elektron ditransfer ke nitrogen cincin nikotinamida yang bermuatan positif, dan atom hidrogen kedua ditransfer ke atom karbon C4 yang berseberangan dengan nitrogen ini. Potensial elektroda standar pasangan redoks NAD+/NADH adalah −0,32 volt, membuat NADH sebagai reduktor kuat. Reaksi ini sangat mudah berbalik arah, ketika NADH direduksi menjadi molekul lain dan dioksidasi kembali menjadi NAD+. Hal ini berarti koenzim ini dapat secara terus-menerus berada dalam siklus NAD+ dan NADH tanpa sendirinya dikonsumsi.
Secara fisik, koenzim ini berbentuk bubuk amorf yang higroskopik dan sangat larut dalam air. Padatan ini stabil jika disimpan dalam keadaan gelap dan kering. Larutan NAD+ tidak berwarna dan stabil selama satu pada temperatur 4 °C dan pH netral. Ia akan terurai dengan cepat apabila terkena asam ataupun basa. Seketika terurai, produk dekomposisi ini merupakan inhibitor enzim.
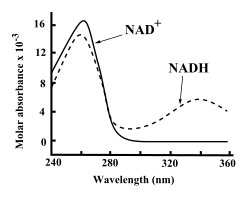
Baik NAD+ dan NADH menyerap ultraviolet dengan sangat kuat oleh karena keberadaan basa adeninanya. Sebagai contoh, puncak absorpsi NAD+ berada pada panjang gelombang 259 nanometer (nm), dengan koefisien pemunahan 16.900 M−1cm−1. NADH juga menyerap panjanga gelombang yang lebih tinggi, dengan puncak kedua dalam absorpsi UV-nya adalah 339 nm dengan koefisien atenuasi molar 6.220 M−1cm−1. Perbedaan spektrum absorpsi ultraviolet antara bentuk koenzim yang teroksidasi dengan yang tereduksi ini membuat zat ini sangat mudah diukur pada uji enzim.
NAD+ dan NADH juga memiliki spektrum fluoresens yang berbeda. NADH dalam larutan memiliki puncak emisi pada 460 nm dan waktu paruh fluoresens sepanjang 0,4 nanosekon, manakala pada koenzim yang teroksidasi tidak memiliki emisi fluoresens. Sifat sinyal fluoresens berubah ketika NADH terikat pada protein, sehingga perubahan ini dapat digunakan untuk mengukur konstanta disosiasi, yang sangat berguna dalam kajian kinetika enzim. Perubahan dalam sinyal fluoresens ini juga digunakan untuk mengukur perubahan dalam keadaan redoks sel hidup, melalui mikroskopi fluoresens.
Konsentrasi dan keadaan zat dalam sel
Dalam hati tikus, kandungan total NAD+ dan NADH adalah kira-kira 1 μmol per gram berat basah hewan, sekitar 10 kali lipat konsentrasi NADP+ dan NADPH dalam sel yang sama. Konsentrasi sebenarnya NAD+ dalam sitosol sel sulit diukur, dengan perkiraan terbaru dalam sel hewan berkisar antara 0,3 M sampai dengan 1,0 sampai 2,0 mM dalam ragi. Namun, sekitar 80% zat ini terikat pada protein, sehingga konsentrasinya dalam larutan haruslah lebih rendah.
Data konsentrasi zat ini pada bagian sel lainnya sangat terbatas, walaupun dalam mitokondria konsentrasi NAD+ sama dengan konsentrasi zat ini dalam sitosol. NAD+ dibawa ke dalam mitokondria melalui protein transpor membran yang khusus oleh karena koenzim ini tidak dapat berdifusi melewati membran.
Keseimbangan antara bentuk NAD yang teroksidasi dengan bentuk yang tereduksi disebut rasio NAD+/NADH. Rasio ini merupakan komponen penting yang disebut sebagai keadaan redoks sel. Keadaan redoks sel adalah ukuran yang mencerminkan baik aktivitas metabolisme sel maupun kesehatan sel. Efek rasio NAD+/NADH terhadap sel sangatlah kompleks. Ia mengontrol aktivitas beberapa enzim kunci, meliputi gliseraldehida 3-fosfat dehidrogenase dan piruvat dehidrogenase. Dalam jaringan mamalia yang sehat, perkiraan rasio NAD+/NADH umumnya berkisar sekitar 700; sehingga lebih menguntungkan bagi reaksi oksidasi. Sebaliknya, rasio NADP+/NADPH umumnya sekitar 0,005.
Biosintesis
NAD+ disintesis melalui dua lintasan metabolisme yaitu lintasan de novo dan lintasan penyelamatan (salvage). Lintasan de novo menggunakan asam amino, sedangkan lintasan penyelamatan dilakukan dengan mendaur ulang komponen-komponen prekursor seperti nikotinamida menjadi NAD+. Lintasan yang lain disebut lintasan kinurenina yang dilalui oleh triptofan. Lintasan kinurenina terbagi dua, yang pertama adalah lintasan asam kinurenat, yang kedua adalah lintasan asam kuinolinat dan hidroksikynurenina-3. Ketiga senyawa organik tersebut merupakan prekursor dari NAD+.
Produksi de novo

Kebanyakan organisme mensintesis NAD+ dari komponen-komponen yang sederhana. Reaksi yang terlibat berbeda-beda dari organisme yang satu ke organisme lain. Namun, terdapat kesamaan dalam penghasilan asam kuinolinat dari asam amino tertentu seperti triptofan pada hewan dan beberapa bakteri, ataupun asam aspartat pada beberapa bakteri dan tumbuhan. Asam kuinolinat diubah menjadi asam nikotinat adenina dinukleotida (NaMN) melalui transfer gugus fosforibosa. Gugus adenilat kemudian ditransfer untuk membentuk asam nikotinat adenina dinukleotida (NaAD). Pada akhirnya, gugus asam nikotinat pada NaAD diamidasi menjadi gugus nikotinamida (Nam), membentuk nikotinamida adenina dinukleotida.
Pada langkah lebih lanjut, beberapa NAD+ diubah menjadi NADP+ oleh NAD+ kinase, yang memfosforilasi NAD+. Pada kebanyakan organisme, enzim ini menggunakan ATP sebagai sumber gugus fosfat, walaupun pada bakteri seperti Mycobacterium tuberculosis dan arkea seperti Pyrococcus horikoshii menggunakan polifosfat anorganik sebagai donor fosfat alternatif.

Lintasan penyelamatan (salvage)
Selain perakitan NAD+ secara de novo menggunakan asam amino sederhana, sel juga mendaur ulang senyawa-senyawa yang mengandung nikotinamida untuk menghasilkan NAD+. Walaupun terdapat banyak prekursor-prekursor yang diketahui, terdapat tiga senyawa alamiah mengandung cincin nikotinamida yang digunakan dalam lintasan penyelamatan (daur ulang) ini, yakni asam nikotinat (Na), nikotinamida (Nam), dan nikotinamida ribosida (NR). Prekursor-prekursor ini kemudian dimasukkan ke dalam lintasan biosintesis NAD(P)+ melalui reaksi adenilasi dan fosforibosilasi seperti yang ditunjukkan pada ilustrasi di atas. Senyawa-senyawa ini dapat berasal dari makanan, di mana campuran asam nikotinat dan nikotinamida disebut sebagai vitamin B3 ataupun niasin. Namun, senyawa-senyawa ini juga dapat diproduksi dalam sel sendiri, yaitu melalui pelepasan gugus nikotinamida dari NAD+ dalam reaksi transfer ADP-ribosa. Enzim-enzim yang terlibat dalam lintasan daur ulang ini tampaknya terkonsentrasi dalam inti sel, yang mengompensasikan laju konsumsi NAD+ yang tinggi dalam organel ini. Sel juga dapat mendapatkan NAD+ secara ekstraseluler (luar sel) dari sekelilingnya.
Walaupun terdapat lintasan de novo, lintasan daur ulang ini merupakan lintasan yang esensial pada manusia. Kekurangan niasin pada makanan mengakibatkan penyakit defisiensi vitamin pelagra. Kebutuhan NAD+ yang tinggi ini disebabkan oleh konsumsinya yang tinggi pada reaksi modifikasi pascatranslasi.
Lintasan daur ulang yang digunakan oleh mikroorganisme berbeda dengan lintasan yang digunakan oleh mamalia. Beberapa patogen seperti ragi Candida glabrata dan bakteri Haemophilus influenzae adalah auksotrof NAD+ (yakni tidak dapat mensintesis NAD+). Namun mereka memiliki lintasan daur ulang, sehingga sangat bergantung pada sumber luar NAD+ dan prekursornya. Bahkan pada patogen seperti Chlamydia trachomatis, ia tidak memiliki gen untuk biosintesis maupun daur ulang NAD+ dan NADP+, sehingga harus menerima asupan koenzim ini dari sel inangnya.
Fungsi

Nikotinamida adenina dinukleotida memiliki beberapa peranan esensial dalam metabolisme. Ia berperan sebagai koenzim pada reaksi redoks, sebagai donor gugus ADP-ribosa pada reaksi ADP-ribosilasi, sebagai prekursor molekul kurir kedua ADP-ribosa siklik, dan juga sebagai substrat bagi enzim DNA ligase bakteri dan enzim sirtuin yang menggunakan NAD+ untuk melepaskan gugus asetil dari protein.
Oksidoreduktase
Peran utama NAD+ dalam metabolisme adalah mentransfer elektron dari satu molekul ke molekul lainnya. Reaksi seperti ini dikatalisasi oleh sekelompok besar enzim yang dinamakan oksidoreduktase. Tata nama enzim dalam kelompok oksidoreduktase mengandung nama kedua substratnya. Sebagai contoh, NADH-ubikuinon oksidoreduktase mengkatalisis oksidasi NADH oleh koenzim Q. Namun, enzim oksidoreduktase ini juga dapat dirujuk sebagai dehidrogenase ataupun reduktase. Biasanya NADH-ubikuinon oksidoreduktase disebut sebagai NADH dehidrogenase ataupun kadang kala koenzim Q reduktase.
Ketika terikat pada suatu protein, NAD+ dan NADH biasanya terikat pada motif struktural yang dikenal dengan nama lipatan Rossmann. Motif ini dinamakan atas nama Michael Rossmann yang merupakan ilmuwan yang pertama kali memperhatikan banyaknya motif ini pada protein pengikat nukleotida. Lipatan ini mengandung tiga atau lebih lempengan beta paralel yang dihubungkan oleh dua heliks alfa dengan urutan beta-alfa-beta-alfa-beta. Oleh karena tiap lipatan Rossmann mengikat satu nukleotida, domain pengikatan untuk dinukleotida NAD+ terdiri dari dua lipatan Rossmann yang berpasangan, dengan tiap lipatan mengikat satu nukleotida. Walau demikian, lipatan ini tidaklah universal ada pada enzim yang bergantung pada NAD. Baru-baru ini ditemukan suatu kelas enzim bakteria yang terlibat dalam metabolisme asam amino mengikat koenzim ini, namun tidak memiliki motif lipatan Rossmann.

Ketika terikat pada tapak aktif suatu oksidoreduktase, cincin nikotinamida koenzim ini diposisikan sedemikiannya ia dapat menerima hidrida dari substrat enzim lainnya. Oleh karena karbon C4 yang menerima hidrogen ini prokiral, hal ini dapat digunakan dalam kinetika enzim untuk mengetahui mekanisme enzim. Hal ini dilakukan dengan mencampurkan enzim dengan substrat yang beratom deuterium sebagai pengganti hidrogen, sehingga enzim akan mereduksi NAD+ dengan mentransfer deuterium daripada hidrogen. Dalam kasus ini, enzim dapat menghasilkan salah satu stereoisomer NADH. Pada beberapa jenis enzim, hidrogen ditransfer dari atas bidang cincin nikotinamida. Enzim demikian disebut sebagai oksidoreduktase kelas A, manakala enzim kelas B mentransfer atom hidrogennya dari bawah bidang.
Walaupun terdapat kemiripan pada cara protein mengikat koenzim NAD+ dan NADP+, enzim hampir selalu memiliki spesifisitas yang tinggi untuk mengikat hanya salah satu dari NAD+ maupun NADP+. Spesifisitas ini mencermikan peranan metabolik kedua koenzim yang berbeda dan merupakan akibat dari perbedaan residu asam amino yang berbeda pada kantong pengikat koenzim tersebut. Sebagai contohnya, pada tapak aktif enzim pengikat NADP, ikatan ion terbentuk antara rantai samping asam amino basa dengan gugus fosfat NADP+ yang asam. Sebaliknya, pada enzim yang mengikat NAD, muatan kantongnya terbalik, menjauhkan NADP+ untuk berikatan dengannya. Walau demikian, terdapat pengecualian terhadap kaidah ini. Enzim seperti aldosa reduktase, glukosa-6-fosfat dehidrogenase, dan metilenatetrahidrofolat reduktase dapat menggunakan kedua enzim tersebut pada beberapa spesies organisme.
Peran dalam reaksi redoks
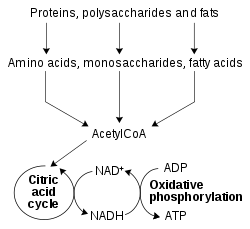
Reaksi redoks melibatkan oksidoreduktase sangat penting dalam metabolisme, terutama untuk membongkar energi yang tersimpan dalam ikatan rangkap oksigen yang relatif lemah. Pada proses ini, senyawa tereduksi seperti glukosa dan asam lemak akan dioksidasi, sehingga melepaskan energi kimia dari O2. Sementara itu, NAD+ direduksi menjadi NADH, sebagai bagian dari oksidasi beta, glikolisis, dan siklus asam sitrat. Pada eukariota, elektron yang dibawa oleh NADH yang diproduksi di sitoplasma ditransfer ke mitokondria (untuk mereduksi NAD+mitokondria) oleh antar-jemput mitokondria, seperti antar-jemput malat-aspartat. NADH mitokondria kemudian dioksidasi oleh rantai transpor elektron, yang memompa proton melintasi membran dan menghasilkan ATP melalui fosforilasi oksidatif. Sistem antar-jemput ini juga memiliki fungsi transportasi yang sama dalam kloroplas.
Karena bentuk teroksidasi dan tereduksi dari nikotinamida adenin dinukleotida digunakan dalam rangkaian reaksi yang terkait ini, sel mempertahankan konsentrasi NAD+ dan NADH yang signifikan, dengan rasio NAD+/NADH yang tinggi memungkinkan koenzim ini bertindak sebagai pengoksidasi dan agen pereduksi. Sebaliknya, fungsi utama NADPH adalah sebagai agen pereduksi dalam anabolisme, dengan koenzim ini terlibat dalam jalur seperti sintesis asam lemak dan fotosintesis. Karena NADPH diperlukan untuk mendorong reaksi redoks sebagai zat pereduksi kuat, rasio NADP+/NADPH dijaga sangat rendah.
Meskipun penting dalam katabolisme, NADH juga digunakan dalam reaksi anabolik, seperti glukoneogenesis. Kebutuhan NADH dalam anabolisme ini menimbulkan masalah bagi prokariota yang tumbuh pada nutrisi yang hanya melepaskan sejumlah kecil energi. Misalnya, bakteri nitrifikasi seperti Nitrobacter mengoksidasi nitrit menjadi nitrat, yang melepaskan energi yang cukup untuk memompa proton dan menghasilkan ATP, tetapi tidak cukup untuk menghasilkan NADH secara langsung. Karena NADH masih diperlukan untuk reaksi anabolik, bakteri ini menggunakan nitrit oksidoreduktase untuk menghasilkan gaya gerak proton yang cukup untuk menjalankan bagian dari rantai transpor elektron secara terbalik dan menghasilkan NADH.
Peran non-redoks
Koenzim NAD+ juga digunakan dalam reaksi transfer ADP-ribosa. Misalnya, enzim yang disebut ADP-ribosiltransferase menambahkan bagian ADP-ribosa dari molekul ini ke protein, dalam modifikasi pascatranslasi yang disebut ADP-ribosilasi. ADP-ribosilasi melibatkan baik penambahan bagian ADP-ribosa tunggal, dalam ribosilasi mono-ADP, maupun transfer ADP-ribosa ke protein dalam rantai bercabang panjang, yang disebut poli(ADP-ribosil)asi. Mono-ADP-ribosilasi pertama kali diidentifikasi sebagai mekanisme sekelompok toksin bakteri, terutama toksin kolera, tetapi juga terlibat dalam pensinyalan sel normal. Poli(ADP-ribosil)asi dilakukan oleh poli(ADP-ribosa) polimerase (PARP). Struktur poli(ADP-ribosa) terlibat dalam regulasi beberapa peristiwa seluler dan paling penting dalam inti sel, dalam proses seperti perbaikan DNA dan pemeliharaan telomer. Selain fungsi-fungsi di dalam sel, sekelompok ADP-ribosiltransferase ekstraseluler baru-baru ini ditemukan, tetapi fungsinya belum diketahui dengan jelas. NAD+ juga dapat ditambahkan ke RNA seluler sebagai modifikasi ujung 5'.

Aksi ekstraseluler dari NAD+
Dalam beberapa tahun terakhir, NAD+ juga telah dikenal sebagai molekul pensinyalan ekstraseluler yang terlibat dalam komunikasi sel-ke-sel. NAD+ dilepaskan dari neuron dalam pembuluh darah,kandung kemih,usus besar, dari sel neurosekretori, dan dari sinaptosom otak, dan diusulkan untuk menjadi neurotransmitter baru yang mengirimkan informasi dari saraf ke sel efektor di organ otot polos. Pada tumbuhan, NAD ekstraseluler menginduksi resistansi terhadap infeksi patogen, dan reseptor NAD ekstraseluler telah diidentifikasi. Penelitian lebih lanjut diperlukan untuk menentukan mekanisme yang mendasari aksi NAD+ ekstraseluler dan pentingnya untuk kesehatan manusia dan proses kehidupan pada organisme lain.
Signifikansi klinis
Enzim yang membuat dan menggunakan NAD+ dan NADH penting dalam farmakologi dan penelitian pengobatan penyakit. Desain dan pengembangan obat memanfaatkan NAD+ dalam tiga cara: i) sebagai target langsung, ii) merancang inhibitor enzim atau aktivator yang mengubah aktivitas enzim yang bergantung pada NAD, iii) menghambat biosintesis NAD+.
NAD+ merupakan target langsung dari obat isoniazid, yang digunakan dalam pengobatan tuberkulosis (disebabkan oleh Mycobacterium tuberculosis). Isoniazid merupakan suatu prodrug, dan setelah memasuki bakteri diaktifkan oleh enzim peroksidase, yang mengoksidasi senyawa menjadi bentuk radikal bebas. Radikal ini kemudian bereaksi dengan NADH, menghasilkan produk aduk yang merupakan inhibitor sangat poten terhadap enzim enoil-asill karier protein reduktase, dan dihidrofolat reduktase.
Banyak oksidoreduktase menggunakan NAD+ dan NADH sebagai substrat, sehingga ditemukannya inhibitor berdasarkan NAD+ yang spesifik untuk satu enzim cukup mengejutkan. Kerja inhibitor melibatkan pengikatan menggunakan motif struktural yang sangat lestari. Sebagai contoh, inhibitor berdasarkan senyawa asam mikofenolat dan tiazofurin menghambat IMP dehidrogenase di tempat pengikatan NAD+. Enzim ini penting dalam metabolisme purin, sehingga senyawa ini berguna sebagai obat antikanker, antivirus, atau imunosupresif. Terdapat obat lain yang bukan penghambat enzim, tetapi mengaktifkan enzim yang terlibat dalam metabolisme NAD+. Sirtuin merupakan target yang sangat menarik, karena aktivasi deasetilasi bergantung pada NAD ini memperpanjang usia pada beberapa model hewan. Senyawa seperti resveratrol meningkatkan aktivitas enzim ini, yang penting dalam kemampuannya untuk menunda penuaan pada organisme model. Dalam satu percobaan, tikus yang diberi NAD selama satu minggu meningkatkan komunikasi antara inti sel dan mitokondria.
Karena perbedaan jalur metabolisme biosintesis NAD+ antara organisme, seperti antara bakteri dan manusia, area metabolisme ini merupakan area yang menjanjikan untuk pengembangan antibiotik baru. Misalnya, enzim nikotinamidase, suatu enzim yang mengubah nikotinamida menjadi asam nikotinat, merupakan target untuk desain obat, karena enzim ini tidak ada pada manusia tetapi ada dalam ragi, bakteri, dan Leishmania.
Dalam bakteriologi, NAD kadang-kadang disebut faktor V, digunakan sebagai suplemen untuk media kultur untuk beberapa bakteri rumit yang memerlukan nutrian khusus.
Sejarah

Koenzim NAD+ pertama kali ditemukan oleh ahli biokimia Inggris Arthur Harden dan William John Young pada 1906. Mereka menunjukkan bahwa menambahkan ekstrak ragi yang direbus dan disaring sangat mempercepat fermentasi alkohol dalam ekstrak ragi yang tidak direbus. Mereka menyebut faktor tak dikenal yang bertanggung jawab atas efek ini sebagai kofermen. Melalui pemurnian dari ekstrak ragi dengan proses yang lama dan sulit , faktor stabil panas ini diidentifikasi sebagai fosfat gula nukleotida oleh Hans von Euler-Chelpin. Pada 1936, ilmuwan Jerman Otto Heinrich Warburg menunjukkan fungsi koenzim nukleotida dalam transfer hidrida dan mengidentifikasi bagian nikotinamida sebagai tempat reaksi redoks.
| Perpustakaan nasional | |
|---|---|
| Lain-lain | |