
| Virus hepatitis B
| |
|---|---|
|
Hepatitis B virus | |
 | |
| Komposisi genom virus ICTV |
double-stranded DNA reverse transcriptase virus (en) |
| Penyakit |
Hepatitis B dan Hepatitis E |
| Taksonomi | |
| Superdomain | Biota |
| Domain | Virus |
| Famili | Hepadnaviridae |
| Genus | Orthohepadnavirus |
| Spesies |
Hepatitis B virus |
Virus hepatitis B (sering disingkat HBV) adalah virus DNA untai ganda, spesies dari genus Orthohepadnavirus, dan anggota dari famili virus Hepadnaviridae. Virus ini menyebabkan penyakit hepatitis B.
Penyakit
Di samping menyebabkan hepatitis, infeksi HBV dapat menyebabkan sirosis hati dan karsinoma hepatoselular.
Virus ini juga telah diduga dapat meningkatkan risiko kanker pankreas.
Peran
Infeksi virus oleh virus hepatitis B (HBV) menyebabkan banyak perubahan hepatosit yang disebabkan oleh aksi langsung dari protein yang dikodekan virus, HBx, dan perubahan tidak langsung yang disebabkan oleh peningkatan drastis spesi oksigen reaktif (ROS) intraseluler setelah infeksi. HBx muncul sebagai disregulat sejumlah jalur seluler. HBx menyebabkan disregulasi sebagian dengan berikatan pada DNA genom, mengubah pola ekspresi miRNA, mempengaruhi histon metiltransferase, berikatan dengan protein SIRT1 untuk mengaktivasi transkripsi, dan berkooperasi dengan histon metilase dan demetilase untuk mengubah pola ekspresi sel. HBx sebagian bertanggung jawab atas peningkatan sekitar 10,000 kali lipat ROS intraselular pada infeksi kronis HBV. Meningkatnya ROS sebagian dapat disebabkan oleh lokalisasi HBx pada mitokondria dengan HBx menurunkan potensial membran mitokondria. Sebagai tambahan, protein HBV lainnya, HBsAg, juga meningkatkan ROS melalui interaksi dengan retikulum endoplasma.
Peningkatan spesi oksigen reaktif (ROS) setelah infeksi HBV menyebabkan inflamasi yang mengakibatkan peningkatan ROS lebih lanjut. ROS menyebabkan lebih dari 20 tipe kerusakan DNA. Kerusakan DNA oksidatif bersifat mutagenik. Lebih lanjut, perbaikan kerusakan DNA dapat menyebabkan perubahan epigenetik pada tempat yang rusak selama perbaikan DNA. Perubahan epigenetik dan mutasi dapat menyebabkan gangguan dalam proses seluler yang kemudian berkontribusi terhadap penyakit hati. Seiring dengan akumulasi perubahan epigenetik dan mutasi akhirnya menyebabkan perkembangan kanker, perubahan epigenetik muncul dengan peran lebih besar dalam karsinogenesis dibandingkan dengan mutasi. Hanya satu atau dua gen, TP53 dan mungkin ARID1A, termutasi pada lebih dari 20% kasus kanker hati sementara 41 gen memiliki promoter yang hipermetilasi (menekan ekspresi gen) pada lebih dari 20% kasus kanker hati, dengan tujuh dari gen ini mengalami hipermetilasi pada lebih dari 75% kasus kanker hati. Lebih lanjut mengenai perubahan pada tempat perbaikan DNA, perubahan epigenetik juga disebabkan oleh HBx merekrut enzim DNA metiltransferase, DNMT1 dan/atau DNMT3A, ke lokus gen spesifik untuk mengubah tingkat metilasi dan ekpresi gen tersebut. HBx juga mengubah asetilasi histon yang dapat berdampak pada ekspresi gen.
Ribuan gen pengkode protein muncul sebagai situs pelekatan HBx. Lebih lanjut, sekitar 15 RNA-Mikro dan 16 RNA bukan pengode panjang juga terdampak oleh pelekatan HBx pada promoternya. Setiap RNA-Mikro yang berubah dapat berdampak pada ekspresi ratusan RNA duta.
Klasifikasi
Virus hepatitis B diklasifikasikan sebagai spesies tipe Orthohepadnavirus, yang memuat tiga spesies lainnya: virus hepatitis tupai tanah, virus hepatitis marmot tanah, dan virus hepatitis B monyet berbulu wol. Genus ini diklasifikasikan sebagai bagian dari famili Hepadnaviridae, yang memuat dua genus lainnya, Avihepadnavirus dan yang kedua sampai sekarang belum ditentukan. Famili virus ini belum ditentukan pada ordo virus. Virus serupa hepatitis B telah ditemukan pada seluruh kera (orang utan, ungka, gorila, dan simpanse), monyet Dunia Baru, dan monyet berbulu wol Dunia Baru yang menimbulkan pemikiran mengenai nenek moyang virus ini pada primata.
Virus ini dibagi menjadi empat serotipe besar (adr, adw, ayr, ayw) berdasarkan keberadaan epitop antigenik pada protein sampul. Serotipe ini didasarkan pada determinan umum (a) dan dua pasang determinan saling taktumpang (d/y dan w/r). Galur virus juga dibagi ke dalam sepuluh genotipe (A–J) dan empat puluh subgenotipe menurut variasi sekuens nukleotida keseluruhan dari genom. Genotipe ini memiliki perbedaan persebaran geografi yang digunakan dalam menelusuri evolusi dan transmisi virus. Perbedaan di antara genotipe berdampak pada keparahan penyakit, penanganan dan kemungkinan komplikasi, serta respons pada pengobatan dan kemungkinan vaksinasi. Serotipe dan genotipe tidak semestinya berhubungan.
Genotipe D memiliki 10 subgenotipe.
Spesies takterklasifikasi
Sejumlah spesies seperti virus hepatitis B takterklasifikasi telah diisolasi dari kelelawar.
Morfologi
Struktur
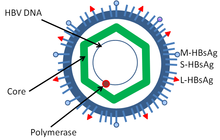
Virus hepatitis B merupakan anggota famili Hepadnaviridae. Partikel virus, disebut partikel Dane (virion), terdiri dari sampul lipid luar dan inti nukleokapsid ikosahedral yang tersusun atas protein. Nukleokapsid melingkupi DNA virus dan DNA polimerase yang memiliki aktivitas serupa transkriptase balik pada retrovirus. Sampul luar memuat protein sematan yang terlibat dalam penempelan virus pada, dan masuk ke dalam, sel yang rentan. Virus ini merupakan salah satu virus hewan bersampul terkecil dengan diameter virion 42 nm, tetapi terdapat bentuk pleomorfik, termasuk badan berfilamen dan berbentuk bola tanpa inti. Partikel ini tidak menularkan penyakit dan tersusun atas lipid dan protein yang merupakan bagian bentuk dari permukaan virion, yang disebut antigen permukaan (HBsAg), dan diproduksi berlebih selama siklus hidup virus.
Komponen
Virus ini terdiri atas:
- HBsAg
- HBcAg (HBeAg merupakan varian sambatan)
- DNA polimerase virus hepatitis B
- HBx (Fungsi protein ini belum diketahui dengan baik, tetapi bukti mengacu pada bagian peran dalam aktivasi proses transkripsi virus.).
Virus hepatitis D membutuhkan partikel sampul HBV untuk menjadi virulen.
Evolusi
Evolusi awal hepatitis B, seperti semua virus, sulit untuk ditentukan.
Divergensi Orthohepadnavirus dan Avihepadnavirus terjadi ~125.000 tahun yang lalu (interval 95% 78.297–313.500). Avihepadnavirus dan Orthohepadnavirus mulai membuat variasi sekitar 25.000 tahun yang lalu. Percabangan saat ini memicu munculnya genotipe A–H Orthohepadnavirus. Galur manusia memiliki nenek moyang bersama termuda dengan penanggalan 7.000 (interval 95%: 5.287–9.270) sampai 10.000 (interval 95%: 6.305–16.681) tahun yang lalu.
Avihepadnavirus tidak memiliki protein X tetapi masih terdapat bingkai pembacaan X vestigial di dalam genom hepadnavirus bebek. Protein X mungkin telah berevolusi dari DNA glikosilase.
Laju mutasi nonsinonim pada virus ini diperkirakan sekitar 2×10−5 penggantian asam amino per situs per tahun. Jumlah rata-rata substitusi nukleotida/situs/tahun adalah ~7.9×10−5.
Perkiraan kedua asal-usul virus ini mengusulkan nenek moyang bersama termuda dari galur manusia berevolusi ~1.500 tahun yang lalu. Nenek moyang bersama termuda dari galur unggas telah berada pada 6.000 tahun yang lalu. Laju mutasi diperkirakan sebesar ~10−6 substitusi/situs/tahun.
Analisis lainnya dengan himpunan data lebih besar mengusulkan bahwa hepatitis B menginfeksi manusia 33.600 tahun yang lalu (kepadatan posterior lebih tinggi 95% 22.000–47.100 tahun yang lalu. Perkiraan laju substitusi sebesar 2.2 × 10−6 substitusi/situs/tahun. Peningkatan populasi signifikan tercatat dalam 5.000 tahun terakhir. Infeksi lintas spesies pada orangutan dan ungka terjadi dalam 6.100 tahun terakhir.
Pemeriksaan sekuens pada finch zebra telah menekan asal-usul genus ini kembali setidaknya pada 40 juta tahun silam dan kemungkinan 80 juta tahun silam. Spesies kluster simpanse, gorila, orangutan, dan ungka dengan manusia terisolasi. Spesies nonprimata termasuk virus hepatitis tikus tanah, virus hepatitis tupai tanah, dan virus hepatitis tupai arktik. Sejumlah spesies penginfeksi kelelawar telah dideskripsikan. Spesies ini diusulkan bahwa spesies kelelawar Dunia Baru mungkin merupakan nenek moyang dari spesies primata.
Sebuah penelitian yang mengisolasi populasi manusia Arktik sirkumpolar telah mengusulkan nenek moyang subgenotipe B5 (tipe endemik yang ditemukan di dalam populasi ini) berasal dari nenek moyang virus di Asia sekitar 2.000 tahun yang lalu (kepadatan posterior lebih tinggi 95% 900 SM – 830 M). Penggabungan terjadi sekitar 1000 M. Subgenotipe ini menyebar dari Asia, bermula dari Greenland dan kemudian menyebar ke barat dalam 400 tahun terakhir.
Genom

Ukuran
Genom HBV terbentuk dari DNA sirkuler, tetapi genomnya tak biasa karena DNA tidak sepenuhnya berunting rangkap. Satu ujung unting panjang penuh terpaut dengan DNA polimerase virus. Genom memiliki panjang 3020–3320 nukleotida (untuk unting panjang penuh) dan 1700–2800 nukleotida (untuk unting pendek).
Pengodean
Unting bermakna negatif (bukan pengode) berkomplementer dengan RNA duta virus. DNA virus ditemukan pada nukleus segera setelah menginveksi sel. DNA berunting rangkap parsial berubah menjadi unting rangkap penuh akibat penyelesaian unting bermakna (+) oleh DNA polimerase sel (DNA polimerase virus digunakan untuk tahap berikutnya) serta penghilangan molekul protein dari unting bermakna (-) dan sekuens RNA pendek dari unting bermakna (+). Basa bukan pengode dihilangkan dari ujung unting bermakna (-) dan ujung tersebut digabung kembali.
Gen virus ditranskripsi oleh RNA polimerase II sel di dalam inti sel dari cetakan DNA sirkuler tertutup secara kovalen (cccDNA). Dua enhanser bernama enhanser I (EnhI) dan enhanser II (EnhII) telah diidentifikasi di dalam genom HBV. Kedua enhanser menampilkan aktivitas yang lebih besar pada sel asal hati, dan bersama mereka menjalankan dan meregulasi ekspresi transkripsi virus penuh. Terdapat empat gen diketahui yang dikodekan oleh genom disebut C, P, S, dan X. Protein inti dikodekan oleh gen C (HBcAg), dan kodon mulainya diawali oleh kodon mulai AUG dalam bingkai menghulu dari tempat protein prainti diproduksi. HBeAg diproduksi oleh proses proteolitik protein prainti. DNA polimerase dikodekan oleh gen P. Gen S merupakan gen yang mengkode antigen permukaan (HBsAg). Gen HBsAg ini memiliki satu bingkai pembacaan terbuka yang panjang, tetapi memuat tiga kodon "mulai" (ATG) di dalam bingkai yang membagi gen ke dalam tiga bagian, pra-S1, pra-S2, dan S. Karena kodon mulai ganda ini, tiga polipeptida dengan ukuran berbeda disebut besar, sedang, dan kecil (pra-S1 + pra-S2 + S, pra-S2 + S, dan S) diproduksi. Fungsi protein yang dikodekan oleh gen X belum sepenuhnya dipahami, tetapi beberapa bukti mengacu bahwa fungsi protein ini sebagai transaktivator transkripsi.
Sejumlah unsur RNA bukan pengode telah diidentifikasi di dalan genom HBV, yaitu: PRE-alfa HBV, PRE-beta HBV, dan RNA sinyal enkapsidasi epsilon HBV.
Genotipe
Perbedaan genotipe setidaknya 8% dari sekuens yang membedakan penyebaran geografi dan perbedaan ini telah dihubungkan dengan sejarah antropologi. Subtipe genotipe telah dideskripsikan, berbeda 4–8% dari genom.
Terdapat delapan genotipe yang diketahui dilabeli A hingga H.
Kemungkinan genotipe baru "I" telah dideskripsikan, tetapi penerimaan notasi ini tidak universal.
Dua genotipe lebih jauh telah dikenali. Daftar saat ini (2014) memuat A hingga J. Beberapa subtipe juga telah dikenali, setidaknya terdapat 24 subtipe.
Genotipe berbeda mungkin merespons pengobatan dengan jalan berbeda.
Genotipe individual
Tipe F yang berdivergen dari genom lainnya sebesar 14% merupakan tipe paling berdivergen yang diketahui. Tipe A pada umumnya ditemukan di Eropa, Afrika, dan Asia Tenggara, termasuk Filipina. Tipe B dan C mendominasi di Asia; tipe D umum ditemukan di wilayah Mediterania, Timur Tengah, dan India; tipe E terlokalisasi di Afrika sub-Sahara; tipe F (atau H) terbatas pada Amerika Selatan dan Tengah. Tipe G telah ditemukan di Prancis dan Jerman. Genotipe A, D, dan F mendominasi Brazil dan semua genotipe terdapat di Amerika Serikat dengan frekuensi bergantung pada etnisitas.
Galur E dan F masing-masing berasal dari populasi suku pedalaman Afrika dan Dunia Baru.
Tipe A memiliki dua subtipe: Aa (A1) di Afrika/Asia dan Filipina serta Ae (A2) di Eropa/Amerika Serikat.
Tipe B memiliki dua persebaran geografis berbeda: Bj/B1 ('j'—Jepang) dan Ba/B2 ('a'—Asia). Tipe Ba selanjutnya terbagi lagi menjadi empat klad (B1–B4).
Tipe C memiliki dua subtipe geogragis: Cs (C1) di Asia Tenggara dan Ce (C2) di Asia Timur. Subtipe C dibagi ke dalam lima klad (C1–C5). Klad keenam (C6) telah terdeskripsi di Filipina, tetapi hanya terdapat di satu isolat sampai saat ini. Tipe C1 diasosiasikan dengan Vietnam, Myanmar, dan Thailand; tipe C2 dengan Jepang, Korea, dan Republik Rakyat Tiongkok; tipe C3 dengan Kaledonia Baru dan Polinesia; C4 dengan Australia; serta C5 dengan Filipina. Subtipe lebih jauh telah terdeskripsi di Papua, Indonesia.
Tipe D terbagi ke dalam 7 subtipe (D1–D7).
Tipe F terbagi ke dalam 4 subtipe (F1–F4). F1 lebih lanjut terbagi menjadi 1a dan 1b. Di Venezuela, subtipe F1, F2, dan F3 ditemukan di Amerindian Timur dan Barat. Di Amerindian Selatan hanya F3 yang ditemukan. Subtipes Ia, III, dan IV menunjukkan persebaran geografi yang terbatas (secara berurutan Amerika Tengah, Utara Amerika Selatan, dan Selatan Amerika Selatan) sementara klad Ib dan II ditemukan pada seluruh Amerika kecuali berturut-turut pada Utara Amerika Selatan dan Amerika Utara.
Siklus hidup
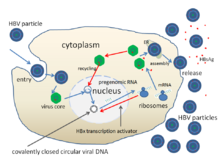
Siklus hidup virus hepatitis B cukup kompleks. Hepatitis B merupakan satu dari beberapa virus nonretroviral diketahui yang menggunakan transkriptase balik sebagai bagian dari proses replikasi.
- Penempelan
- Virus memperoleh pintu masuk ke dalam sel dengan berikatan ke reseptor pada permukaan sel dan masuk dengan endositosis dimediasi oleh klatrin atau kaveolin-1. HBV pada awalnya berikatan ke proteoglikan heparin sulfat. Segmen pra-S1 dari protein L HBV kemudian berikatan dengan kuat ke reseptor permukaan sel polipeptida kotranspor natrium taurokolat (NTCP), dikodekan oleh gen SLC10A1. NTCP umumnya ditemukan pada membran sinusoidal sel hati. Keberadaan NTCP pada sel hati berkorelasi dengan spesifisitas jaringan infeksi HBV.
- Penetrasi
- Setelah endositosis, membran virus berfusi dengan membran sel inang, melepaskan nukleokapsid ke dalam sitoplasma.
- Pembukaan
- Karena virus memperbanyak melalui RNA yang dibuat oleh enzim sel inang, DNA genom virus dikirim menuju inti sel. Proses ini terjadi dengan mentranspor kapsid pada mikrotubulus ke pori inti. Protein inti berdisosiasi dari DNA virus berunting rangkap parsial, yang kemudian dibuat unting rangkap penuh (oleh DNA polimerase inang) dan bertransformasi menjadi DNA sirkuler tertutup secara kovalen (cccDNA) yang menyediakan cetakan untuk transkripsi empat RNA duta virus.
- Replikasi
- RNA duta terbesar, lebih panjang dari genom virus, digunakan untuk membuat salinan genom baru dan membuat protein inti kapsid serta DNA-polimerase-bergantung-RNA-virus.
- Perakitan
- Empat transkrip virus ini melalui proses tambahan dan bergabung untuk membentuk virion turunan yang kemudian dilepaskan dari sel atau kembali menuju inti dan membentuk siklus kembali untuk memproduksi lebih banyak salinan.
- Pelepasan
- RNA duta panjang kemudian ditranspor kembali ke sitoplasma dengan virion protein P menyintesis DNA melalui aktivitas transkriptase balik.
Transaktivasi gen
HBV memiliki kemampuan untuk mentransaktivasi FAM46A.